
ads:

UNIVERSIDADE FEDERAL DE UBERLÂNDIA
FACULDADE DE ENGENHARIA ELÉTRICA
PÓS-GRADUAÇÃO EM ENGENHARIA ELÉTRICA
Modelos de Predição Linear para a Análise
de Sinais Eletroencefalográficos (EEG) e
de Matrizes Multieletrodo (MEA)
Jaqueline Alves Ribeiro
Maio
2006
ads:

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.
UNIVERSIDADE FEDERAL DE UBERLÂNDIA
FACULDADE DE ENGENHARIA ELÉTRICA
PÓS-GRADUAÇÃO EM ENGENHARIA ELÉTRICA
Modelos de Predição Linear para a Análise
de Sinais Eletroencefalográficos (EEG) e
de Matrizes Multieletrodo (MEA)
Jaqueline Alves Ribeiro
Texto da dissertação apresentada à Universidade Federal de
Uberlândia, perante a banca de examinadores abaixo, como parte dos
requisitos necessários para a obtenção do título de Mestre em Ciências.
Aprovada em 29 de maio de 2006.
Banca Examinadora:
Prof. Elisângela P. Silveira Lacerda, Dra. (ICB/UFG)
Prof. Gilberto Arantes Carrijo, Dr. (FEELT/UFU)
Prof. João Batista Destro Filho, Dr. - Orientador (FEELT/UFU)
Prof. João Yoshiyuki Ishihara, Dr. (ENE/UnB)
Prof. Ubirajara Coutinho Filho, Dr. (Fequi/UFU)
Prof. Wilson Felipe Pereira, Dr. (ICBIM/UFU)
ads:

FICHA CATALOGRÁFICA
Elaborada pelo Sistema de Bibliotecas da UFU / Setor de Catalogação e Classificação
R482m
Ribeiro, Jaqueline Alves, 1981-
Modelos de predição linear para a análise de sinais
eletroencefalográficos (EEG) e de matrizes multieletrodo (MEA)
/ Jaqueline Alves Ribeiro. - Uberlândia, 2006.
174 f. : il.
Orientador: João Batista Destro Filho.
Dissertação (mestrado) - Universidade Federal de Uberlândia,
Programa de Pós-Graduação em Engenharia Elétrica.
Inclui bibliografia.
1. Engenharia biomédica - Teses. 2. Processamento de sinais - Teses. I.
Destro Filho, João Batista. II. Universidade Federal de Uberlândia. Programa
de Pós-Graduação em Engenharia Elétrica. III. Título.
CDU: 61:62

Modelos de Predição Linear para a Análise
de Sinais Eletroencefalográficos (EEG) e
de Matrizes Multieletrodo (MEA)
Jaqueline Alves Ribeiro
Texto da dissertação apresentada à Universidade Federal de Uberlândia como parte dos
requisitos para obtenção do título de Mestre em Ciências.
Prof. João Batista Destro Filho, Dr.
Orientador
Prof. Darizon Alves de Andrade, Ph.D.
Coordenador do curso de Pós-Graduação
Aos meus pais, Antônio e Luzia,
as minhas irmãs Adriana e Andrea
e ao meu afilhado Gabriel Victor
pelo apoio e incentivo.
Esta não é apenas uma conquista minha,
mas sim de todos vocês, por tudo que fizeram por mim.
Obrigada!
iv
Agradecimentos
A Deus pela vida e por guiar meus caminhos, por todas as bênçãos concedidas
para superar os momentos difíceis nesta jornada. Minha fé em Ti me fez acreditar que as
vitórias surgem através de esforços.
Aos meus amados pais, Antônio e Luzia, pelos princípios com que me criaram,
carinho e amor que tenho recebido; as minhas irmãs, Adriana e Andrea, em especial à
Andrea por me ajudar a superar a dor nos momentos mais difíceis, pela amizade, e
sempre estar do meu lado, incondicionalmente. Ao meu sobrinho Gabriel Victor, o
filho da dindinha, que me faz ver a vida do jeito dele, com as brincadeiras e risadas; ao
meu cunhado Alessandro, pelo apoio prestado, às vindas em Uberlândia, às mudanças,
e a toda minha família, tios, tias, avós, primos e primas, que sempre torceram pelo
meu sucesso.
À madrinha Ondina por todo carinho e apoio prestado durante esta caminhada,
seus conselhos e sua amizade. Suas filhas Lara e Laís, pelas descontrações.
À amiga Flávia, minha mãezinha de Uberlândia, pelo carinho, força e
dedicação, por estar sempre por perto me auxiliando, em particular, às idas ao
supermercado, à hidroginástica, à fisioterapia, e aos amigos que conheci a partir dela, à
célula Geração Águia pelos ensinamentos bíblicos. Foi muito gratificante conhecer
vocês!
Aos meus amigos de longe e de perto pela força e inspiração nessa jornada, as
conversas “jogadas fora”, as brincadeiras, as comidas comunitárias na Kitnet, os
conselhos, os passeios, os puxões de orelha merecidos, enfim... Só Deus sabe o quanto
v
foram importantes quando os colocaram em meu caminho. Obrigada por fazerem parte
da minha vida!
Ao meu orientador João Batista Destro Filho, pelo constante estímulo e toda
atenção dedicada, e também pela compreensão nas minhas ausências, por toda
confiança depositada em mim para a finalização da dissertação.
Aos coordenadores da Pós-graduação com os quais tive contato durante minha
caminhada pela compreensão e apoio, acreditando em minha capacidade para concluir
esta jornada; à secretária Marli Junqueira Buzzi pela dedicação e carinho e a todos os
funcionários da Faculdade de Engenharia Elétrica da Universidade Federal de
Uberlândia que contribuíram de alguma maneira para a conclusão desta dissertação.
À CAPES pela oportunidade do financiamento, mas que não foi possível até o
final, pois tive que me ausentar durante o tratamento.
Ao Laboratório de Engenharia Biomédica (BIOLAB) pelo amparo
tecnológico e estrutural, aos colegas do BIOLAB, pela amizade e apoio durante esta
jornada, em especial, àqueles que conviveram comigo, Ailton Jr., Eduardo, Geovane,
Guilherme, Fábio, Karina, Marcelle, Saulo, Tiago Finotti. Obrigada a todos pelas
contribuições!!
A todos os professores que compõem a banca, pelas discussões realizadas para a
finalização deste trabalho, e também a disponibilidade em participar da defesa.
Aos alunos de Iniciação Científica, Rodrigo R. Cardoso e Aline R. de Assis
pela ajuda no processamento dos sinais, com esforço e dedicação. Ao acadêmico de
Graduação de Engenharia Elétrica (UFU) Ronaldo J. Viana, por ter estudado toda a
base de dados EEG que compõem este trabalho.
À Universidade de Tampere (Finlândia), em particular ao Prof. A. Värri, pelo
fornecimento da base de dados EEG. Também ao Prof. S. Martinoia, ao doutorando A.
N. Ide e todos os colegas da DIBE – Universidade de Gênova, Itália, pelo fornecimento
dos dados MEA utilizados neste trabalho. À Prof. Nathalia Peixoto, Universidade de
Washington, USA, pelo registro das atividades elétricas neurais em microcrustáceos.
vi
Ao Dr. Aguinaldo Bertucci (Hospital de Clínicas de Uberlândia),
neurocirurgião, e ao doutorando Fábio J. Parreira (BIOLAB), pelo intenso trabalho de
análise e classificação do banco de dados EEG.
Ao Prof. Rodrigo Lemos Pinto (UFG) pelas referências bibliográficas, quando
estava em Goiânia.
Aos colegas da Pós-Graduação, pelo companheirismo, colaboração e
aprendizagem que desenvolvemos juntos.
Por fim, agradeço a todos que direta e indiretamente contribuíram para a
realização desse feito, por me auxiliarem na imensa e inacabável tarefa de auto-
conhecimento.
vii
Não sei...
Se a vida é curta ou longa demais pra nós,
Mas sei que nada do que vivemos tem sentido,
Se não tocamos o coração das pessoas.
Muitas vezes basta ser:
Colo que acolhe,
Braço que envolve,
Palavra que conforta,
Silêncio que respeita,
Alegria que contagia,
Lágrima que corre,
Olhar que acaricia,
Desejo que sacia,
Amor que promove.
E isso não é coisa de outro mundo, é o que dá sentido à vida.
É o que faz com que ela não seja nem curta, nem longa demais,
Mas que seja intensa, verdadeira, pura...
Enquanto durar...
Feliz aquele que transfere o que sabe e aprende o que ensina.”
Cora Coralina
viii
Resumo
RIBEIRO, J. A. Modelos de Predição Linear para análise de Sinais
Eletroencefalográficos (EEG) e de Matrizes Multieletrodo (MEA). Uberlândia: FEELT-
UFU, 2006, 174 f.
Esta dissertação objetiva propor modelos de sinais neurofisiológicos, oriundos
de matrizes multieletrodo (MEA), utilizadas para a aquisição da atividade neural
espontânea a partir de células em cultura; bem como medidas neurológicas obtidas de
eletroencefalografia (EEG). Os modelos são desenvolvidos no contexto aplicativo de
MEAs utilizadas como neuroimplantes, com especial enfoque na detecção e prevenção
de crises epilépticas a partir de sinais EEG ou de medidas mais invasivas, obtidas em
nível celular. Sob este enfoque, as técnicas de processamento de sinais devem satisfazer
um compromisso complicado, exigindo baixa complexidade computacional e operação
em tempo real. Tais restrições levam à escolha da filtragem linear adaptativa, baseada
em modelo auto-regressivo, e à teoria de estatísticas de ordem elevada (EOE) para,
respectivamente, processar sinais não-estacionários e considerar a não-linearidade dos
sistemas envolvidos. O filtro de predição linear funcionou adequadamente para ambos
os sinais, sendo que para o caso da MEA, ele foi mais estável e preciso. Observou-se
um menor tempo de convergência para sinais EEG relativamente aos sinais MEA, os
quais, em sua maioria, podem ser considerados não-gaussianos e correlacionados,
ocorrendo uma atividade periódica em eletrodos vizinhos. A atividade neural
espontânea de pequenos grupos de neurônios pode ser considerada um ruído branco.
ix
Palavras-chave: predição linear, estatística de ordem elevada,
eletroencefalografia, matriz multieletrodo, neuroimplante, codificação neural.
x
Abstract
RIBEIRO, J. A. Linear-Prediction Models for Electroencephalographic (EEG) and
Multielectrode-Array (MEA) Signal Analysis. Uberlândia: FEELT-UFU, 2006, 174 f.
This work establishes models of neurophysiological signals, which are
composed of spontaneous activity measurements taken by means of multielectrode
arrays (MEAs) applied on in vitro cell cultures; as well as of neurological signals based
on electroencephalography. These models suppose that MEAs are employed as
neuroprostheses applied for detection and forecast of epileptic seizures, based on EEG
signals or on invasive measurements which are taken in a cellular level. From this point
of view, the signal processing tools must fulfil a problematic trade-off involving low
computational complexity and real-time operation. Such requirements lead to the choice
of auto-regressive adaptive-linear filtering and high-order statistics (HOE) as the
techniques to be used in order to cope with, respectively, non-stationary signals and
nonlinear systems. Linear prediction of both signals is quite efficient, particularly in the
case of MEA signals, for which the model is stable and accurate. On the other hand, the
convergence times for EEG signals are lower then their respective counterparts for
MEA signals, which may be considered mainly non-Gaussian and correlated. Cyclic
activity was also observed for MEA signals associated with neighboring electrodes,
whereas signals recorded from small groups of neurons present a white-noise behaviour.
Key-words: linear prediction, high-order statistics, electroencephalography,
multielectrode arrays, neuroprostheses, neural coding.
xi
Conteúdo
Lista de Figuras................................................................................ XVI
Lista de Tabelas...............................................................................XXII
Lista de Abreviaturas e Símbolos................................................ XXIV
1. Introdução .......................................................................................26
2. Conceitos básicos em neurofisiologia e neurologia .....................31
2.1 Introdução.................................................................................................... 31
2.2 O neurônio .................................................................................................. 32
2.2.1 Membrana Celular............................................................................ 34
2.2.2 Canais iônicos................................................................................... 36
2.3 Sinapses entre neurônios ............................................................................. 39
2.4 Potencial de membrana dos neurônios ........................................................ 42
2.4.1 Potencial de ação.............................................................................. 42
2.4.2 Potencial Pós-Sináptico (PPS).......................................................... 46
2.5 Medidas Neurofisiológicas: Matrizes Multieletrodo (MEA) ...................... 47
2.5.1 Culturas Celulares ............................................................................ 47
2.5.2 Interface bioeletrônica...................................................................... 49
2.5.3 Matrizes Multieletrodo..................................................................... 50
2.5.4 Utilização e Fabricação das MEAs................................................... 52
xii
2.5.5 Histórico ........................................................................................... 55
2.5.6 Aplicações ........................................................................................ 56
2.6 O Sistema Nervoso (SN) ............................................................................. 62
2.6.1 Sistema Nervoso Central .................................................................. 64
2.6.1.1 A medula.................................................................................................. 65
2.6.1.2 O encéfalo................................................................................................ 67
2.6.2 Sistema Nervoso Periférico .............................................................. 70
2.6.2.1 Sistema nervoso Autônomo. .................................................................... 70
2.6.3 Cérebro ............................................................................................. 71
2.6.3.1 O córtex cerebral...................................................................................... 72
2.7 Medidas Neurológicas: O Eletroencefalograma (EEG) .............................. 74
2.7.1 Introdução......................................................................................... 74
2.7.2 Medição............................................................................................ 76
2.7.3 Características estatísticas do sinal EEG.......................................... 80
2.7.4 Tipos de ondas EEG ......................................................................... 81
2.8 Epilepsia ...................................................................................................... 83
2.9 Conclusão .................................................................................................... 85
3. Síntese Bibliográfica sobre Processamento de Sinais EEG e
MEA .................................................................................................88
3.1 Introdução.................................................................................................... 88
3.2 Considerações gerais e processamento digital de sinais EEG..................... 88
3.3 Processamento de sinais MEA .................................................................... 90
3.3.1 Pré-processamento e geração da série temporal e intervalos entre
spikes............................................................................................... 90
3.3.2 Classificação dos spikes.................................................................. 93
xiii
3.3.3 Estimação da conectividade neural ................................................. 95
3.4 Conclusão .................................................................................................... 97
4. Predição Linear Adaptativa e Estatística de Ordem Elevada
(EOE) .............................................................................................100
4.1 Introdução.................................................................................................. 100
4.2 Filtragem Adaptativa................................................................................. 101
4.3 Cálculo dos Filtros .................................................................................... 105
4.3.1 Filtro Ótimo: aproximações de Wiener .......................................... 105
4.3.2 Filtros subótimos: aproximação estocástica .................................. 109
4.4 Predição Linear Direta (“forward”)........................................................... 111
4.4.1 Modelo auto-Regressivo (AR) ....................................................... 112
4.4.2 Modelo da Predição Linear Direta ................................................. 112
4.4.3 Critérios para uma boa predição..................................................... 113
4.4.4 Solução ótima de Wiener para Predição Linear Direta .................. 113
4.4.5 Solução subótima usando aproximação estocástica do Gradiente . 117
4.5 Predição Linear Reversa (“backward”) .................................................... 119
4.5.1 Modelo auto-Regressivo (AR) Reverso ......................................... 119
4.5.2 Modelo da Predição Linear Reversa............................................... 119
4.5.3 Critérios para uma boa predição..................................................... 120
4.5.4 Solução ótima de Wiener para Predição Linear Reversa ............... 120
4.5.5 Solução subótima usando aproximação estocástica do Gradiente
Reversa ........................................................................................... 121
4.6 Estatísticas de Ordem Elevada (EOE)....................................................... 122
4.6.1 Funções com uma variável aleatória ............................................. 123
4.6.2 Funções com várias variáveis aleatórias......................................... 125
xiv
4.6.3 O ruído branco e os índices de descorrelação ................................ 128
4.7 Conclusão .................................................................................................. 129
5. Simulações e Resultados...............................................................132
5.1 Introdução.................................................................................................. 132
5.2 Aquisição dos Sinais ................................................................................. 133
5.3 Ferramentas e Metodologia....................................................................... 135
5.3.1 Processamento dos dados e critérios utilizados na Predição Linear135
5.3.2 Processamento de dados utilizado para estimação dos índices de
descorrelação .................................................................................. 140
5.4 Resultados 1: EEG e Predição Linear ....................................................... 142
5.4.1 Processamento do banco de dados contendo sinais EEG com crise142
5.4.2 Processamento do banco de dados contendo sinais EEG sem crise144
5.4.3 Comparação dos resultados do banco de dados EEG/Crise e
EEG/SemCrise................................................................................ 146
5.5 Resultados 2: MEA e Predição Linear ...................................................... 147
5.5.1 Processamento de dados de sinais MEA – Teste 1......................... 147
5.5.2 Processamento de dados de sinais MEA – Teste 2......................... 150
5.5.3 Resumo dos resultados dos Testes 1 e 2 e comparação dos resultados
obtidos com os sinais EEG............................................................. 151
5.6 Resultados 3: MEA como Ruído Branco .................................................. 153
5.7 Conclusão .................................................................................................. 155
6. Conclusões e Trabalhos Futuros .................................................158
6.1 Conclusões gerais...................................................................................... 158
6.2 Trabalhos futuros....................................................................................... 162
Referências Bibliográficas.................................................................163
xv
Anexos .................................................................................................174
xvi
Lista de Figuras
Figura 1.1 Neurônios reais (adaptada de (LENT, 2001).......................................... 27
Figura 2.1 Um neurônio motor e suas partes constituintes (adaptada de (URL 1)). 33
Figura 2.2 Neurônios em conexão e o botão sináptico no detalhe (adaptada de
(URL 2)) ................................................................................................. 34
Figura 2.3 Desenho esquemático de uma célula, evidenciando suas organelas e a
membrana plasmática (adaptada de (URL 4))........................................ 35
Figura 2.4 Desenhos esquemáticos da membrana plasmática com seus canais
protéicos. Acima uma membrana com seus vários canais numa visão
tridimensional (adaptada de (CARDOSO, 2005)). Abaixo os canais
iônicos em evidência (adaptada de (LENT, 2001))................................ 37
Figura 2.5 Sinapse química (adaptada de (URL 5)) ................................................ 39
Figura 2.6 Botões sinápticos em um neurônio receptor. Eles existem tanto no
soma como nos dendritos, às vezes ocorrendo também no início do
axônio (cone axonal) (adaptada de (GUYTON, 2002)) ......................... 40
Figura 2.7 Botão sináptico em detalhe, evidenciando as vesículas e as
mitocôndrias. Na fenda sináptica estão os neurotransmissores
(adaptada de (FRISÓN, 1999))............................................................... 41
Figura 2.8 Potencial de ação exibido em um osciloscópio (adaptada de (URL 6)). 43
Figura 2.9 Representação de uma onda de despolarização e repolarização
associada ao potencial de ação (adaptada de (GUYTON, 2002)).......... 43
xvii
Figura 2.10 Potencial de ação (adaptada de (VILELA, 2005)) ................................. 44
Figura 2.11 Princípio do “tudo-ou-nada”: em A), o potencial de ação (PA) do
neurônio pré-sináptico gera um potencial pós-sináptico excitatório
(PPSE) que não chega a atingir o limiar; em B), um trem de PAs gera
um PPSE que ultrapassa o valor de limiar, o que faz com que seja
deflagrado um PA no neurônio pós-sináptico (adaptada de (LENT,
2001)) ..................................................................................................... 45
Figura 2.12 Eventos eletroquímicos da sinapse excitatória e sinapse inibitória
(adaptada de (GUYTON, 2002))............................................................ 47
Figura 2.13 Interface bioeletrônica (adaptada de (FROMHERZ, 2003)).................. 49
Figura 2.14 Caminho neurônio- chip - neurônio (adaptada de (FROMHERZ,
2003)). Vide arranjo (c) da Figura 2.13.................................................. 50
Figura 2.15 Conjunto de eletrodos em um tecido nervoso (adaptada de
(RENNAKER, 2005)) ............................................................................ 51
Figura 2.16 Exemplos de aplicação das matrizes de microeletrodos (adaptada de
PEIXOTO, 2001) (a) corte de cultura em tecido do cerebelo sobre
MEA, (b) cultura celular de neurônios isolados do gânglio da raiz
dorsal de ratos em cultura (14 dias)........................................................ 52
Figura 2.17 Matriz tridimensional fabricada com técnicas de
microeletromecanismos (adaptada de (BAY, 2000)) ............................. 53
Figura 2.18 Esquema geral da fabricação da MEA (adaptada de (PEIXOTO, 2001))53
Figura 2.19 (a) Matriz de microeletrodos com neurônios e sinais extracelulares
medidos de vários neurônios em cada eletrodo. Gânglio abdominal de
Aplysia (adaptada de (STENGER, 1994)). (b) Sinais de uma matriz
extracelular versus sinais medidos com eletrodos intracelulares
(adaptada de (BEADLE, 1988))............................................................. 58
Figura 2.20 MEA funcionando como neuroimplante em um nervo fibular (adaptada
de (RUTTEN, 1999)).............................................................................. 59
xviii
Figura 2.21 Esquema de um neuroimplante inteligente (adaptada de (PEREIRA,
2004)) ..................................................................................................... 60
Figura 2.22 Células gliares: astrócitos (nutrição), oligodentrócitos (sustentação) e
células microgliais (defesa) (adaptada de (VILELA, 2005)................... 63
Figura 2.23 Figura esquemática mostrando o SNC, representado pelo encéfalo e
pela medula e o SNP, representado pelos nervos (adaptada de
(FRISÓN, 1999)).................................................................................... 64
Figura 2.24 Meninges: Dura-máter (mais externa), Aracnóide (Intermediária) e
Pia-máter (mais interna)(adaptada de (CARDOSO, 2005))................... 65
Figura 2.25 Medula Espinhal e Encéfalo (adaptada de (CARDOSO, 2005)) ........... 66
Figura 2.26 O encéfalo (adaptada de (URL 6)) ......................................................... 67
Figura 2.27 Diencéfalo e Tronco Encefálico. Vista posterior do encéfalo com
exclusão do cerebelo (adaptada de (VILELA, 2005))............................ 68
Figura 2.28 Ponte e Bulbo em evidência, com o cerebelo ao fundo e parte do
telencéfalo acima. Vista anterior do encéfalo (adaptada de CARDOSO,
2005)) ..................................................................................................... 69
Figura 2.29 Secção mostrando a metade direita do encéfalo vista por dentro,
evidenciando suas subdivisões (adaptada de (VILELA, 2005)) ............ 69
Figura 2.30 Sistema Nervoso Central e Periférico. Evidência de parte do Sistema
Nervoso Autônomo (Simpático e Parassimpático), bem como de
exemplo de nervos motores e sensoriais (adaptada de (CARDOSO,
2005)) ..................................................................................................... 71
Figura 2.31 Lobos do córtex cerebral (adaptada de (VILELA, 2005)) ..................... 72
Figura 2.32 Subdivisões funcionais corticais, cerebelo e tronco encefálico
(adaptada de (VILELA, 2005)) .............................................................. 73
Figura 2.33 Eletrodos posicionados no escalpo e o registro eletroencefalográfico
(adaptada de (LENT, 2001))................................................................... 76
Figura 2.34 Eletrodos de disco de prata (modelos da Grass®) (adaptada de
(BUTTON, 2000)).................................................................................. 77
xix
Figura 2.35 Eletrodos de agulha de platina (modelos da Grass®) (adaptada de
(BUTTON, 2000)).................................................................................. 77
Figura 2.36 Eletrodos nasofaríngeo (adaptada de (BUTTON, 2000))....................... 78
Figura 2.37 Eletrodos prefixados em grade (adaptada de (BUTTON, 2000)). Touca
com eletrodos posicionados (adaptada de (CARDOSO, 2005)) ............ 78
Figura 2.38 Eletrodos tipo clipe de orelha (“ear clip”) (adaptada de (BUTTON,
2000)) ..................................................................................................... 79
Figura 2.39 Eletrodos corticais (adaptada de (BUTTON, 2000)).............................. 79
Figura 2.40 Exemplos de ondas cerebrais (adaptada de (BUTTON, 2000) &
BERNARDI, 1999))............................................................................... 83
Figura 3.1 Forma de onda extracelular com diferentes potenciais de ação (adaptada
de (LEWICKI, 1998)) ............................................................................ 90
Figura 3.2 Métodos de extração da população dos spikes (PS) (adaptada de (CHAN,
2004)) ..................................................................................................... 93
Figura 3.3 Histograma de intervalo de neurônio cruzado. Eixo y: intervalo entre
spikes [ms]. Eixo x: tempo [ms] (adaptada de (CASTELLONE, 2003))96
Figura 4.1 Diagrama de blocos representando o problema de filtragem estatística
(adaptada de (WIDROW, 1985)) ......................................................... 102
Figura 4.2 Filtro Adaptativo usado para a identificação de um sistema (adaptada
de (HAYKIN, 1991))............................................................................ 103
Figura 4.3 Aplicação da filtragem adaptativa ao modelamento inverso (adaptada
de (HAYKIN, 1991))............................................................................ 104
Figura 4.4 Filtragem adaptativa aplicada à predição. A saída 1 corresponde à
predição do valor de entrada; A saída 2 refere-se ao erro de predição
( adaptada de (HAYKIN, 1991)).......................................................... 104
Figura 4.5 Filtro adaptativo utilizado no cancelamento de interferências (adaptada
de (HAYKIN, 1991))............................................................................ 105
xx
Figura 4.6 Ilustração do problema geral do Filtro de Wiener. Dados dois
processos estacionários, x(n) e d(n), que são estatisticamente
descorrelacionados entre si, o filtro W(z) minimiza a estimativa do erro
médio quadrático, d (n), de d(n). (adaptada de (DE FATTA, 1988)). 106
ˆ
Figura 4.7 Filtro Transversal com N coeficientes (adaptada de (HAYKIN, 1991))107
Figura 4.8 Representação gráfica de w1 x freqüência ........................................... 116
Figura 4.9 Representação gráfica de w2 x freqüência ........................................... 116
Figura 4.10 Gráfico da Função Laplaciana, com seus momentos e cumulantes de
ordem 1 a 4 ........................................................................................... 127
Figura 4.11 Gráfico da Função Gaussiana, com seus momentos e cumulantes de
ordem 1 a 4. .......................................................................................... 127
Figura 4.12 Gráfico da Função Uniforme, com seus momentos e cumulantes de
ordem 1 a 4 ........................................................................................... 127
Figura 5.1 Potenciais de ação associados a neurônios do gânglio cerebral do
caramujo Tritonia diomedea (adaptada de (PEIXOTO, 2002)) ........... 134
Figura 5.2 Exemplo de um gráfico típico de eqm x n. Eixo x: tempo. Eixo y: erro
quadrático médio................................................................................... 137
Figura 5.3 Exemplo de EQMr, em um gráfico típico de eqm x n. O valor de EQMr,
calculado como média entre n=400 e n=600, é .................... 138
68≅EQMr
Figura 5.4 Exemplo de um gráfico típico de eqm x n em que o PM representa o
ponto mais alto do gráfico, ocorrendo em n ≈ 600. ............................. 138
Figura 5.5 Exemplo de um gráfico típico de eqm x n em que o TC representa a
convergência do sinal MEA TC
≅
600. ................................................ 139
Figura 5.6 Exemplo de gráfico típico de eqm x n para o caso de um sinal EEG
com crise TE
600
≅
............................................................................... 140
Figura 5.7 Gráfico PM x N para o processamento do banco de dados EEG/Crise.142
xxi
Figura 5.8 Gráfico EQMr x N para o processamento do banco de dados
EEG/Crise............................................................................................. 142
Figura 5.9 Gráfico TC x N para o processamento do banco de dados EEG/
Crise. .................................................................................................... 143
Figura 5.10 Gráfico TE x N para o processamento do banco de dados EEG/Crise. 143
Figura 5.11 Gráfico PM x N para o processamento do banco de dados EEG/Sem
Crise...................................................................................................... 144
Figura 5.12 Gráfico EQMr x N para o processamento do banco de dados EEG/Sem
Crise...................................................................................................... 145
Figura 5.13 Gráfico TC x N para o processamento do banco de dados EEG/Sem
Crise...................................................................................................... 145
Figura 5.14 Gráfico PM x N para o sinal de MEA. ................................................. 148
Figura 5.15 Gráfico EQMr x N para o sinal de MEA.............................................. 148
Figura 5.16 Gráfico TC x N para sinal de MEA...................................................... 149
Figura 5.17 Gráfico PM x N para o sinal de MEA. ................................................. 150
Figura 5.18 Gráfico EQMr x N para o sinal de MEA.............................................. 150
Figura 5.19 Gráfico TC x N para sinal de MEA...................................................... 151
xxii
Lista de Tabelas
Tabela 2.1 Ondas cerebrais (adaptada de (BUTTON, 2000)) ................................. 81
Tabela 4.1 Algumas aplicações da filtragem adaptativa. ....................................... 103
Tabela 4.2 Função Laplaciana................................................................................ 127
Tabela 4.3 Função Gaussiana................................................................................. 127
Tabela 4.4 Função Uniforme.................................................................................. 127
Tabela 4.5 Sumário das variáveis do filtro de Wiener (adaptada de (HAYKIN,
1991)) ................................................................................................... 130
Tabela 5.1 Classificação dos sinais baseados nos índices de descorrelação.......... 141
Tabela 5.2 Comparação dos resultados EEG/Crise e EEG/Sem Crise: Ordem de
filtro e passo de adaptação.................................................................... 146
Tabela 5.3 Comparação dos resultados EEG/Crise e EEG/Sem Crise: Tempo de
convergência e tempo de estacionariedade........................................... 146
Tabela 5.4 Comparação dos resultados EEG/Crise e EEG/Sem Crise: Pico Máximo
e erro quadrático médio de regime permanente. .................................. 147
Tabela 5.5 Comparação dos Resultados MEA: Ordem de filtro e passo de
adaptação .............................................................................................. 152
Tabela 5.6 Comparação dos Resultados MEA: Pico Máximo e erro quadrático
médio de regime permanente................................................................ 152
Tabela 5.7 Comparação dos Resultados MEA: Tempo de convergência .............. 152
xxiii
Tabela 5.8 Índices de descorrelação para diversos sinais ...................................... 153
Tabela 5.9 IDE para os sinais MEA descorrelacionados ....................................... 154
Tabela 5.10 IDE para os sinais MEA correlacionados............................................. 154
Tabela 5.11 IDE para os sinais MEA cíclicos.......................................................... 155
xxiv
Lista de Abreviaturas e Símbolos
ADP Adenosine Diphosphate (Difosfato de Adenosina ou Adenosina di-fosfato)
Ag-AgCl Prata-cloreto de prata
ATP Adenosine triphosphate (Trifosfato de Adenosina ou Adenosia tri-fosfato)
ANE Atividade Neural Espontânea
AR Modelo Auto-Regressivo
CMRR Common-Mode Rejection Ratio (Razão de Rejeição em Modo Comum)
EEG Eletroencefalografia, Eletroencefalograma ou Eletroencefalográfico(a)
eqm Erro Quadrado Médio
EOE Estatística de Ordem Elevada
EQMr Erro Quadrado Médio de Regime Permanente
ERG Eletroretinograma
GPS Global Positioning System (Sistema de Posicionamento Global)
ICM Interface Cérebro-Máquina
IDE Índice de Descorrelação Elevada
K
+
Íon Potássio
LMS Least Mean Square (Mínimos Quadrados Médios)
MEA Multielectrode Array (Matriz Multieletrodo)
N Ordem do Filtro
xxv
Na
+
Íon Sódio
P Íon Fosfato
PA Potencial de Ação
pdf Função de densidade de probabilidade
PDS Processamento Digital de Sinais
PM Pico Máximo
PPS Potencial Pós-Sináptico
PPSE Potencial Pós-sináptico Excitatório
PPSI Potencial Pós-sináptico Inibitório
SN Sistema Nervoso
SNC Sistema Nervoso Central
SNP Sistema Nervoso Periférico
SNR Signal-To-Noise Ratio (Relação Sinal-Ruído)
SUS Sistema Público de Saúde
TC Tempo de Convergência
TE Tempo de Estacionariedade
URL Uniform Resource Locator (localização de informação na Internet)
USA Estados Unidos da América
UTI Unidade de Tratamento Intensivo
μ Passo de Adaptação
Ω Ohms
26
Capítulo 1
Introdução
O cérebro é um órgão que surpreende e impressiona. É como uma ponte, por
assim dizer, entre consciência e mundo exterior. Por isso mesmo tem sido, ao longo
da história humana, objeto de inúmeros questionamentos, instigando cientistas e
filósofos a elaborarem diferentes perspectivas a respeito de suas funções. Todavia,
graças ao avanço dos instrumentos médicos e aos estudos fisiológicos do corpo
humano, o cérebro está sendo melhor compreendido.
Além de funções motoras e sensitivas, o cérebro é responsável ainda pelo
desempenho de importantes atividades superiores como a memória e a cognição.
Trata-se de um dos mais misteriosos órgãos do corpo humano. E por mais que
tentemos desvendar seus segredos, é difícil compreendê-lo.
No entanto, a ciência médica, assim como a engenharia eletrônica, a
computação, e, atualmente, a engenharia neural, seguem de mãos dadas com os
avanços tecnológicos no intuito de melhor conhecer esta máquina, a qual, por sua vez
nos permite conhecer o mundo. Logo, o enfoque das pesquisas envolve o neurônio
(vide Figura 1.1), a unidade básica do sistema nervoso, e o grande responsável pelo
processamento das informações no encéfalo.

27
Figura 1.1 – Neurônios reais (adaptada de (LENT, 2001)).
O desenvolvimento de diversas técnicas, no século XX e no início do século
XXI, capazes de realizar diferentes abordagens de estudo sobre o Sistema Nervoso
Central (SNC), vem possibilitando uma melhor compreensão do seu funcionamento
(HOLLAND, 2003). Contudo, ainda há muitos questionamentos acerca de diversas
patologias que afetam o SNC. Tornou-se possível estudar as atividades cerebrais,
desde uma atividade motora ou sensitiva a uma atividade cognitiva, ou ainda o efeito
de uma patologia. Todavia, diversas disfunções do corpo humano consistem em
desafios para a medicina (Proc. IEEE, 2001). Dentre elas, devem-se destacar a
epilepsia, o controle da dor, e as distrofias. Esses desafios envolvem o
desenvolvimento de modelos matemáticos eficientes e compatíveis ao contexto
fisiológico humano.
Uma das formas de modelar o cérebro se baseia no estudo dos potenciais
elétricos evocados do córtex cerebral e captados por eletrodos dispostos no escalpo.
Para tal estudo, a eletroencefalografia (EEG) consiste em uma importante ferramenta
que permite o diagnóstico e a análise de diversas patologias que perturbam o cérebro,
além do monitoramento de pacientes em enfermarias ou UTIs e o acompanhamento
de fenômenos cognitivos.
Evidentemente, dentre estas patologias, pode-se citar a epilepsia, que atinge
cerca de 1% da população mundial, ou 50 milhões de pessoas, dos quais 25% não
respondem ao tratamento com o método tradicional (URL 10), que preconiza o uso de
28
drogas anticonvulsivantes. Pode-se citar, também, a dor crônica associada a processos
metastáticos em pacientes terminais (Proc. IEEE, 2001).
As novas hipóteses sobre os mecanismos desencadeantes das crises
epilépticas, fruto concreto da interação entre médicos e engenheiros, aliada à
moderna tecnologia de instrumentação biomédica, sugerem novas formas de terapia.
Em LITT e colaboradores (2003), os autores descrevem planos para a construção de
neuroimplantes que minimizariam crises epilépticas, incluindo experimentos
preliminares de eletroestimulação em animais. Tais circuitos funcionariam como
marcapassos cardíacos, monitorando a atividade elétrica do cérebro e identificando
fatores responsáveis por uma crise. Uma vez detectada a possibilidade iminente de
crise, o dispositivo provocaria uma eletroestimulação no cérebro ou em um nervo
periférico, com objetivo de eliminar a crise. Similarmente, neuroimplantes também
poderiam ser utilizados para minimizar sensações dolorosas, através da eletro-
estimulação, ou da neuromodulação obtida através da filtragem seletiva de potenciais
de ação do sistema nociceptivo.
Por outro lado, as recentes pesquisas estão causando uma revolução nos
conceitos de instrumentação neurofisiológica. Dentre elas, estão a interface
bioeletrônica, que permite mecanismos de transdução de um impulso nervoso de um
neurônio para dentro de um circuito eletrônico e vice-versa. Deve-se citar, também, as
matrizes multieletrodo (RUTTEN, 2002), que permitem a aquisição sistemática de
potenciais de ação, utilizando os neurônios em cultura.
Além disso, através de estímulos elétricos e da análise da resposta neural celular
a estes, obtém-se resultados interessantes, diretamente aplicáveis em neurociência e em
procedimentos ligados à reabilitação. Esta pesquisa constitui a base essencial para a
utilização clínica de neuroimplantes especializados e/ou eletroestimulação em seres
humanos, as quais são atualmente limitadas pelos elevados custos da tecnologia de
matrizes multieletrodo (MEA).
De qualquer forma, o desenvolvimento tecnológico da MEA, aliada ao
rompimento de barreiras no modelamento de sistemas complexos, tende em médio e
longo prazos estabelecer novos pontos de vista sobre as patologias do sistema nervoso,
incluindo também os sinais EEG.
29
Enfim, surgem os desafios para a concretização destas novas terapias, que são
paralelos ao desafio do modelamento dos fenômenos biológicos associados. Dentre
estes, para o processamento de sinais, podem-se citar:
9 Propor modelos simples para compreensão intuitiva e desempenho eficiente,
através de algoritmos de baixa complexidade computacional;
9 Estabelecer um conhecimento sobre as características estatísticas do sinal MEA,
buscando novos pontos de vista sobre as patologias que envolvem a alteração da
neurodinâmica, associada à transmissão de informações através do SNC;
9 Processar simultaneamente, através de MEA, vários tipos de sinais biológicos,
tanto em nível celular, como em nível neurológico (EEG).
Esta dissertação representa um passo inicial para o desenvolvimento de
tecnologia própria, em resposta aos desafios mencionados no parágrafo anterior. As
estratégias escolhidas para atingir as metas acima citadas estão descritas em detalhes em
cada um dos capítulos deste trabalho.
No Capítulo 2, faz-se uma síntese dos conceitos básicos de neurofisiologia e
neurologia, bem como sobre as respectivas medidas que fornecem os sinais elétricos
utilizados nesta dissertação: a instrumentação através de MEAs, baseadas em culturas in
vitro, e os exames EEG. Discutem-se também aspectos fisiológicos e epidemiológicos
da epilepsia.
No Capítulo 3, faz-se uma breve síntese das principais técnicas atualmente
empregadas para o processamento de sinais oriundos de EEG e MEA, com enfoque
neste último.
Com isso, no Capítulo 4, apresentam-se as ferramentas teóricas propostas que
tentam, de certa maneira, minimizar as limitações das técnicas atualmente utilizadas na
análise da codificação neural e da epilepsia, buscando atender ao máximo os desafios
especificados como metas deste trabalho.
No Capítulo 5, apresentam-se os resultados obtidos através da aplicação das
técnicas apresentadas no Capítulo 4 à análise de sinais MEA e EEG. Apresentam-se os
cálculos e os resultados finais que conduzem aos modelos simples, em termos das
aproximações auto-regressiva e ruído branco.
30
Por fim, no Capítulo 6, são analisados os resultados obtidos em comparação aos
desafios iniciais. Sugestões importantes para trabalhos futuros, que poderão ser
realizados a partir desta dissertação, são também apresentadas.
As principais contribuições desta dissertação são listadas logo abaixo:
9 A apresentação de um texto único, reunindo informações fisiológicas em nível
celular e neurológico, que propicia uma conexão entre a epilepsia, o EEG, a
MEA e os fenômenos fisiológicos associados (Capítulo 2);
9 A discussão a respeito dos índices de descorrelação elevada (Capítulo 4);
9 O estabelecimento de modelos paramétricos AR para sinais EEG e MEA, de
baixa complexidade computacional (Capítulo 5), permitindo conclusões
importantes no contexto aplicativo de neuroimplantes;
9 A investigação detalhada, até a ordem nº4, sobre a estrutura estatística dos sinais
MEA (Capítulo 5), que sugere novas perspectivas para o estudo da codificação
neural.
31
Capítulo 2
Conceitos básicos em neurofisiologia e
neurologia
2.1 Introdução
Neste capítulo apresentam-se os conceitos sobre os quais se baseiam as medidas
eletrofisiológicas que contextualizam a aplicação deste trabalho. Também se apresentam
as estruturas biológicas de interesse, com suas respectivas instrumentações, em dois
níveis: celular e neurológico.
Inicialmente, faz-se um estudo do neurônio, em particular de suas propriedades
especiais no que diz respeito aos potenciais elétricos. Em seguida, analisa-se o sinal de
matriz de multieletrodo (MEA), sua fabricação e aplicações, como também as novas
técnicas de instrumentação neurofisiológica que sugerem a implementação de
neuroimplantes especializados.
Em seguida, de forma sucinta, explicam-se os princípios gerais do
funcionamento do sistema nervoso, destacando-se algumas áreas funcionais do cérebro,
uma vez que sua compreensão influencia a interpretação dos sinais de
eletroencefalografia (EEG). Consequentemente, faz-se uma breve revisão sobre os
sinais de EEG e suas características estatísticas; e também da epilepsia, suas causas,
tipos e conseqüências na população mundial, particularmente no Brasil.
32
2.2 O neurônio
Todos os estímulos do nosso ambiente, causando sensações como dor e calor; os
sentimentos, pensamentos, programação de respostas emocionais e motoras; a
aprendizagem e memória, a ação de drogas psicoativas, os distúrbios mentais; bem
como qualquer outra ação ou sensação do ser humano, não podem ser entendidas sem o
conhecimento do processo de comunicação entre os neurônios.
O neurônio é uma célula como qualquer outra: possui organelas, núcleo
individualizado, necessita de oxigênio e nutrientes, e é envolta por uma membrana
plasmática. O que diferencia um tipo de célula de outra é a sua função no nosso
organismo. Portanto o que difere os neurônios de outras células, além da sua maior
necessidade metabólica, é a sua função especializada. Ela tem como função receber,
processar e transmitir impulsos eletroquímicos, ou impulsos nervosos, que são sinais de
comunicação e codificação. Pode, assim, ser considerado como a unidade funcional
fundamental do sistema nervoso (LENT, 2001).
Cada neurônio compreende (vide Figura 2.1):
9 um corpo celular que contém um núcleo, retículo endoplasmático muito
abundante (corpos de Nissl), mitocôndrias, aparelho de Golgi e neurofibrilas;
9 uma ou mais ramificações de filamentos citoplasmáticos finos, denominados
dendritos, que conduzem os impulsos até ao corpo celular;
9 um prolongamento, o axônio, que pode ser muito longo e apresentar
ramificações na sua parte distal ou, ao longo da sua extensão, formando
ramificações colaterais. Em alguns casos, os axônios podem estar rodeados por
uma substância esbranquiçada, de natureza lipídica, a mielina, recoberta por
uma película de citoplasma contendo núcleos, designada por bainha de
Schwann. A bainha de mielina é descontínua, dando origem à formação de
nódulos de Ranvier.

33
Figura 2.1 – Um neurônio motor e suas partes constituintes (adaptada de
(URL 1))
Os neurônios são células grandes, com numerosas projeções, denominadas
dendritos. A maioria dos sinais é recebida pelos neurônios em seus dendritos, outros
sinais são recebidos no corpo celular e alguns poucos no início do axônio. Acredita-se
que no cérebro existam cerca de 100 bilhões de neurônios.
Qualquer função cognitiva, como pensar, mover, dormir, olhar, sentir, envolve a
integração de um número desconhecido de neurônios, em áreas específicas do cérebro; e
de estruturas nervosas do organismo, fora do cérebro. Os neurônios se interconectam em
complexas cadeias, e a mensagem viaja através de cada neurônio na forma de impulsos.
Os sinais elétricos transmitidos pelos neurônios se baseiam na movimentação de
íons (átomos, ou grupos de átomos, que perderam ou receberem elétrons), com cargas
positivas ou negativas, que se formam ao longo do neurônio como fruto de reações
químicas. Os neurônios podem ser classificados como:
9 Neurônios sensoriais ou aferentes: transmitem impulsos do exterior para
o sistema nervoso central;
9 Neurônios motores ou eferentes: transmitem impulsos do sistema
nervoso central para o exterior;
9 Neurônios de associação: conduzem impulsos entre os outros dois tipos
de neurônios.
Como qualquer outra célula, o neurônio possui um certo potencial de membrana,
a ser discutido mais adiante. O neurônio, por ser uma célula excitável, tem este
potencial modificado dependendo dos estímulos que sofre através de sinapses (Figura

34
2.2), ou de outros estímulos. A variação do potencial de membrana percorre o axônio
como uma onda de despolarização e repolarização, que vai até os botões sinápticos.
Figura 2.2 – Neurônios em conexão e o botão sináptico no detalhe (adaptada de (URL 2))
Devido à bainha de mielina, a propagação do sinal ocorre de nódulo em nódulo,
uma vez que o potencial de ação somente pode ser regenerado nos nódulos, o que faz
com que este possa chegar ao seu destino de forma mais rápida. Este tipo de propagação
é denominado de condução saltatória. Além disso, a resultante do somatório de
capacitâncias e resistências adicionais das bainhas de mielina, envolvendo o axônio
neuronal, contribuem muito para o aumento da velocidade de condução das fibras
nervosas.
2.2.1 Membrana celular
A membrana neuronal funciona como uma barreira para delimitar o citoplasma e
excluir certas substâncias presentes que banham os neurônios. Sua espessura é de 5 nm,
aproximadamente, e está repleta de proteínas e lipídeos. A função dos neurônios não
pode ser compreendida sem o conhecimento da estrutura e função da sua membrana,
bem como das proteínas associadas.
Quase 70% de toda massa corpórea de um ser humano adulto é basicamente
composto de água, sendo o resto composto principalmente por sais dissolvidos na água,
lipídios, proteínas e glicose. Logo, pode-se dizer que toda célula pode ser considerada
como uma cápsula contendo uma solução aquosa. O que separa esta cápsula do meio
externo, denominado meio extracelular, é uma membrana semipermeável constituída

35
basicamente de lipídios e proteínas (membrana lipoprotéica), conhecida como
membrana plasmática (Figura 2.3).
Quando um sal se dissolve em água, suas ligações iônicas se desfazem, uma vez
que a elevada constante dielétrica da água diminui a intensidade da força iônica e as
moléculas de água solvatem os íons. Ou seja, sais dissolvidos em água se transformam
em íons livres (cátions e ânions, moléculas eletricamente carregadas positivamente e
negativamente), o que origina potenciais elétricos tanto no meio intra quanto no meio
extracelular.
Figura 2.3 – Desenho esquemático de uma célula, evidenciando suas organelas e a membrana
plasmática (adaptada de (CARDOSO, 2005))
Devido a este fenômeno, e como a composição iônica do meio intracelular se
difere da composição do meio extracelular, as células do nosso corpo apresentam uma
diferença de potencial elétrico entre o meio interno e o meio externo, que é denominada
de potencial de membrana ou potencial de repouso da membrana. O interior da
membrana celular apresenta uma carga elétrica menor que a carga elétrica exterior.
Portanto, o potencial elétrico interno é negativo em relação ao potencial elétrico
externo, que é tomado por referência.
A origem desse potencial elétrico pode ser explicada a partir da compreensão
dos fenômenos eletroquímicos protagonizados pela membrana plasmática e por dois
importantes íons existentes em nosso corpo, dentro e fora das células: sódio (Na
+
) e
potássio (K
+
).
Quando em equilíbrio (dinâmico), existe uma diferença de concentração desses
íons entre os meios intra e extracelular: há maior concentração de Na
+
no meio externo
à célula e de K
+
no meio interno, além disso, há maior concentração de ânions orgânicos
dentro da célula. Isso acontece devido à semipermeabilidade da membrana plasmática e
36
devido à sua capacidade de selecionar o trânsito de substâncias entre a célula e o meio
em que ela se encontra, denominada permeabilidade seletiva.
A água trafega de um lado ao outro da membrana celular por pressão osmótica.
Ou seja, apesar de ser altamente insolúvel nos lipídios da membrana, a água atravessa
prontamente a totalidade da membrana celular, passando, quase toda ela, através dos
canais existentes nas moléculas protéicas (GUYTON, 2002).
Podemos afirmar que o canal protéico se liga a uma molécula de ATP (trifosfato
de adenosina), que trafega no meio intracelular. Ocorre então a quebra, formando ADP
(difosfato de adenosina) e P (fosfato), liberando energia. Assim, o canal se modifica,
atrai e se liga a três íons de sódio (três cargas positivas) no lado interno da membrana e
a dois íons de potássio (duas cargas positivas) do lado externo. Portanto, injeta o
potássio e ejeta o sódio. Isso ocorre de forma mais rápida que o funcionamento dos
canais de transporte passivo, o que contribui para a permanência de um potencial de
cerca de -70 mV no meio intracelular em relação ao meio extracelular. Esse processo é
denominado de equilíbrio dinâmico.
Além de contribuir para a manutenção do potencial de membrana num dado
valor de tensão, uma das principais funções da bomba Na
+
/ K
+
é o controle do volume
das células. Sem o funcionamento dessa bomba, a maioria das células do corpo sofreria
tumefação, até ocorrer sua ruptura (GUYTON, 2002).
2.2.2 Canais iônicos
Os canais iônicos regulam o fluxo de íons através da membrana de todas as
células. Em particular, nas células neurais e musculares, eles são importantes para o
controle das rápidas variações do potencial de membrana, associadas ao potencial de
ação e aos potenciais pós-sinápticos das células-alvo (KANDEL, 2000).
Esses canais são grandes proteínas que atravessam toda a estrutura da
membrana, incluindo grupos carboidratos (glicoproteínas) presos à sua superfície,
conforme a Figura 2.4. Todos os canais apresentam um poro aquoso central, que se
estende de uma face à outra da membrana. Muitos canais são formados por duas ou
mais subunidades, que podem ser idênticas ou diferentes, permitindo o fluxo de íons de

37
modo seletivo, continuamente ou em resposta a estímulos elétricos, químicos ou
mecânicos (LENT, 2001).
Figura 2.4 – Desenhos esquemáticos da membrana plasmática com seus
canais protéicos. Acima uma membrana com seus vários canais numa visão
tridimensional (adaptada de (CARDOSO, 2005)). Abaixo os canais iônicos
em evidência (adaptada de (LENT, 2001))
A membrana plasmática de todas as células, inclusive das neurais, consiste em
um mosaico de lipídios e de proteínas. A superfície dessa membrana é formada por uma
dupla camada de fosfolipídios. Nessa lâmina lipídica contínua, ficam embebidas as
moléculas de proteína, inclusive os canais iônicos.
A facilidade com que um íon se desloca dentro de uma solução (sua
mobilidade ou constante de difusão) não depende simplesmente da sua dimensão, como
também do tamanho da molécula do solvente. O canal iônico abre e fecha de forma
tudo-ou-nada, resultando em breves pulsos de corrente através da membrana. Se o
potencial elétrico variar através da membrana, a corrente que flui pelo canal se altera
38
proporcionalmente. A corrente é linearmente relacionada à força propulsora; em outras
palavras, o canal se comporta como um resistor elétrico (KANDEL, 2000).
Os canais iônicos são classificados em dois tipos:
9 os canais catiônicos, que, na maioria das vezes, permitem a passagem de
íons sódio, quando estão abertos; mas às vezes, também permitem a
passagem de íons potássio e/ou cálcio;
9 os canais aniônicos, que permitem, sobretudo, a passagem de íons
cloreto, mas, também, de quantidades diminutas de outros ânions
(GUYTON, 2002).
Os canais catiônicos que conduzem íons sódio são revestidos por cargas
negativas. Essas cargas atraem os íons sódio carregados positivamente para dentro do
canal, quando o diâmetro deste aumenta até um tamanho maior que aquele do íon sódio
hidratado. Essas mesmas cargas negativas, porém, repelem os íons cloreto e outros
ânions, impedindo sua passagem.
Já para os canais aniônicos, quando seu diâmetro se torna suficientemente
grande, conduzem íons cloreto para seu interior e seguem para o lado oposto; enquanto
os cátions sódio, potássio e cálcio são bloqueados, porque a dimensão de seus íons
hidratados impedem sua passagem pelos canais aniônicos.
Portanto, uma substância transmissora que abre os canais catiônicos é
denominada de transmissor excitatório. Ao contrário, a abertura de canais aniônicos,
permitindo a entrada de cargas elétricas negativas, inibe o neurônio. Essas substâncias
transmissoras que abrem estes canais são denominadas de transmissores inibitórios.
Quando a substância transmissora ativa o canal iônico, este se abre em fração de
milissegundos; quando a substância transmissora não está mais presente, o canal se
fecha de modo igualmente rápido. Desta forma, a abertura e o fechamento dos canais
iônicos permite um meio para o controle rápido dos neurônios.

39
2.3 Sinapses entre neurônios
Os neurônios devem desencadear informações sobre o estado interno do
organismo e seu ambiente externo, avaliar esta informação, e coordenar atividades
apropriadas à situação e às necessidades atuais do organismo. Cada neurônio se
comunica com o seguinte por meio de sinapses nervosas (Figura 2.5), que pode ser
definida como o conjunto de fenômenos bioquímicos ocorrendo no espaço de conexão
entre os neurônios. O número de conexões de um único neurônio pode variar de
algumas unidades até algumas centenas de milhares, segundo (GUYTON, 2002).
Figura 2.5 - Sinapse química (adaptada de (URL 3))
A célula nervosa cuja terminação do axônio se conecta ao soma ou ao dendrito
do outro neurônio é denominado de neurônio pré-sináptico, sendo responsável pelo
envio do impulso. O neurônio seguinte, que recebe o impulso, é denominado de
neurônio pós-sináptico. Entre esses dois neurônios existe a fenda sináptica, de largura
compreendendo 200 a 300 Angstroms.
Aproximadamente 80% a 95% dos botões ou terminais pré-sinápticos (Figura
2.6) se situam nos dendritos dos neurônios pós-sinápticos, e apenas 5% a 20% se situam
sobre o soma (GUYTON, 2002).

40
Figura 2.6 – Botões sinápticos em um neurônio receptor. Eles existem tanto
no soma como nos dendritos, às vezes ocorrendo também no início do axônio
(cone axonal) (adaptada de (GUYTON, 2002))
São as sinapses que possibilitam a transmissão do impulso nervoso de um
neurônio ao outro, ou de um neurônio a uma fibra muscular, determinando, inclusive, o
sentido que este impulso deve tomar. Tal impulso é a transmissão de um sinal
codificado, como resultado de um estímulo ocorrido ao longo da membrana do
neurônio. Dois tipos de fenômenos estão envolvidos no processamento do impulso
nervoso: elétricos e químicos. Eventos elétricos propagam um sinal dentro do neurônio,
e processos químicos transmitem o sinal de um neurônio a outro ou a uma célula
muscular. Os processos químicos ocorrem no final do axônio, que libera substâncias
químicas ou neurotransmissores, os quais se unem a receptores químicos situados nos
canais iônicos da membrana do neurônio seguinte, permitindo assim a troca de
informações entre duas células nervosas.
Por exemplo, um neurônio motor colinérgico cujo neurotransmisssor principal é
a acetilcolina, e que inerva uma célula muscular esquelética, produzirá uma ação
sináptica excitatória nesta célula. Já um neurônio motor, também colinérgico, que inerva

41
uma célula muscular cardíaca, produzirá uma ação sináptica inibitória (LENT, 2001),
permitindo que a ação do neurônio pós-sináptico seja restringida ou estimulada.
As sinapses químicas são extremamente importantes porque permitem a
condução do impulso nervoso numa única direção: o impulso segue do neurônio pré-
sináptico (transmissor) ao neurônio pós-sináptico (receptor). A importância dessa
transmissão unidirecional se justifica pela necessidade do sinal ser enviado a
determinadas áreas especificas do sistema nervoso, bem como pela necessidade de
atuação de controle em determinados grupos musculares, ou em glândulas secretoras.
No terminal pré-sináptico, temos duas estruturas bastante importantes: as
vesículas sinápticas, que contém as substâncias neurotransmissoras a serem liberadas na
fenda sináptica; e as mitocôndrias, que fornecem energia (ATP – trifosfato de
adenosina) para a síntese desses mesmos neurotransmissores (Figura 2.7). Estes últimos
apresentam ação rápida. Cada vez que um potencial de ação atinge o terminal pré-
sináptico, algumas vesículas liberam, ao mesmo tempo, o transmissor para o interior da
fenda sináptica, dentro de um intervalo de 1 ms ou menos. A ação desses transmissores
sobre os canais iônicos da membrana do neurônio pós-sináptico também ocorre dentro
de 1 ms ou menos.
Figura 2.7 – Botão sináptico em detalhe, evidenciando as vesículas e as
mitocôndrias. Na fenda sináptica estão os neurotransmissores (adaptada de
(FRISÓN, 1999))
42
2.4 Potencial de membrana dos neurônios
A membrana do neurônio apresenta uma propriedade muito particular que o
distingue da maioria das células do organismo. Essa propriedade, a excitabilidade,
permite que o neurônio produza e transmita a outros neurônios os sinais elétricos em
código, que constituem a linguagem do sistema nervoso (LENT, 2001).
Os biopotenciais encefálicos podem ser diferenciados em dois tipos (BUTTON,
2000): o Potencial de Ação e o Potencial Pós-Sináptico (PPS). O primeiro caracteriza a
transmissão dos impulsos eletroquímicos pelos axônios, ocorrendo de forma assíncrona
e em direções ortogonais à superfície do escalpo. O segundo é o potencial de membrana
resultante no neurônio pós-sináptico, com amplitude muito reduzida (LENT, 2001).
2.4.1 Potencial de ação
Os pesquisadores britânicos Alan Hodgkin (1914-1998) e Andrew Huxley
(1917) foram os pioneiros em estudos para desvendar a bioeletrogênese do impulso
elétrico do neurônio (LENT, 2001). Na conversão entre a energia bioelétrica, que ocorre
na sinapse, os potenciais de ação convergem para o terminal sináptico, e a energia
química, representada pela quantidade de neurotransmissor, é liberada na fenda
sináptica.
O potencial de ação é uma alteração rápida e brusca da diferença de potencial
transmembrana. A membrana do neurônio é polarizada em repouso em torno de -70
mV. O potencial de ação consiste de uma redução rápida da negatividade da membrana
até 0 mV e inversão deste potencial até valores de aproximadamente de +30 mV,
seguido de um retorno também rápido até valores negativos entre zero e o potencial de
repouso, -70mV, conforme a Figura 2.8.

43
Figura 2.8 - Potencial de ação exibido em um osciloscópio (adaptada de (URL 4))
O potencial de ação é um fenômeno de natureza eletroquímica e ocorre devido a
modificações na permeabilidade da membrana do neurônio, que permitem a passagem
de íons de um lado para o outro da membrana. Como os íons são partículas carregadas
eletricamente, ocorrem também modificações no campo elétrico gerado por essas
cargas.
A membrana de alguns tipos de neurônios é polarizada em repouso com valor
cerca de -90 mV (GUYTON, 2002). Com a despolarização, há um pico de até cerca de
+35 mV. Observa-se um aumento rápido do potencial de membrana; seguido de uma
descida também rápida, até um valor inferior ao valor de repouso (cerca de -100 mV), e,
por fim, um lento retorno até o valor de repouso de -90 mV (Figura 2.9).
Figura 2.9 – Representação de uma onda de despolarização e repolarização
associada ao potencial de ação (adaptada de (GUYTON, 2002))
No entanto, para que seja deflagrado um potencial de ação ao longo do axônio, é
necessário que o estímulo sofrido pela membrana do neurônio (no dendrito, no corpo,
ou no cone axonal) seja capaz de elevar abruptamente o potencial da membrana de 15 a

44
30 mV (GUYTON, 2002), ou seja, o potencial deve chegar a cerca de -65 mV
(potencial de limiar).
Apesar do impulso nervoso ocorrer no sentido do axônio, a onda de
despolarização/repolarização da membrana se propaga em todos os sentidos a partir do
ponto em que foi deflagrada (Figura 2.10). Todavia esta onda cessa ao atingir a região
da membrana próxima ao corpo do neurônio, bem como aos dendritos. Com relação ao
axônio, a onda segue até os botões sinápticos e lá desencadeia uma reação química, que
caracterizará a comunicação do neurônio com a célula seguinte.
Em virtude do gradiente de concentração e da carga negativa do fluido
extracelular, os íons entram na fibra através dos canais iônicos. A entrada de sódio
despolariza a membrana, isto é, a face da membrana imersa no fluido extracelular das
fibras se torna menos negativo, em relação ao interior. Se esta despolarização,
denominada potencial gerador, alcança o nível crítico (limiar), a membrana irá gerar um
potencial de ação.
Figura 2.10 – Potencial de ação (adaptada de (VILELA, 2005))

45
Uma vez atingido o limiar, o potencial de ação ocorre com amplitude e duração
fixas. Se o limiar não for atingido, ou seja, a despolarização ou o influxo de sódio não
forem suficientemente fortes, não ocorre o potencial de ação (Figura 2.11). Este
fenômeno é conhecido como princípio do tudo-ou-nada (GUYTON, 2002).
Ocasionalmente, o potencial de ação alcança um ponto na membrana onde não
gerará voltagem suficiente para estimular a área adjacente da membrana. Quando isso
ocorre, a propagação da despolarização é interrompida. Portanto, para que a propagação
de um impulso continue, a proporção entre o potencial de ação e o limiar de excitação
deve ser, a qualquer tempo, maior que 1.
Figura 2.11 – Princípio do “tudo-ou-nada”: em A), o potencial de ação (PA)
do neurônio pré-sináptico gera um potencial pós-sináptico excitatório
(PPSE) que não chega a atingir o limiar; em B), um trem de PAs gera um
PPSE que ultrapassa o valor de limiar, o que faz com que seja deflagrado
um PA no neurônio pós-sináptico (adaptada de (LENT, 2001))
À medida que se abrem os canais de sódio, abrem-se também os canais de
potássio, entretanto de forma mais lenta. Quando os canais de sódio estão
completamente abertos, os de potássio ainda estão se abrindo, ou seja, com o passar do
tempo, aumenta a quantidade de potássio que sai da célula, fazendo com que a
amplitude do potencial de ação comece a diminuir. Quanto menos positivo o potencial
de membrana, menos sódio entra na célula, e com a saída de mais potássio, mais o
potencial avança no sentido negativo, retornando ao repouso.
46
Porém, antes da membrana se repolarizar, o potencial ultrapassa o potencial de
repouso, devido ainda à lentidão dos canais de potássio, que estão se fechando
novamente. Desta forma, o potencial vai lentamente retornando ao valor de repouso.
Isso pode ser observado na Figura 2.9.
2.4.2 Potencial Pós-Sináptico (PPS)
Para uma sinapse excitatória, o resultado do potencial de ação do neurônio pré-
sináptico é a geração de um PPS despolarizante, ou seja, um potencial que tende a
aproximar-se do potencial de limiar. Isso faz com que o neurônio pós-sináptico tenha
mais facilidade em disparar potenciais de ação.
Quando sinapses excitatórias descarregam sobre as superfícies dos dendritos ou
do soma de um neurônio, gera-se um potencial pós-sináptico que persiste por muitos
milissegundos, especialmente quando alguma das substâncias transmissoras sinápticas
de longa duração está envolvida. Esse potencial pode continuar a excitar o neurônio,
fazendo com que ele transmita uma salva de impulsos de saída (GUYTON, 2002).
Já no caso de uma sinapse inibitória, o resultado do potencial de ação do
neurônio pré-sináptico é a geração, no neurônio seguinte, de um PPS hiperpolarizante,
ou seja, um potencial que tende a afastar-se do potencial de limiar. Isso faz com que o
neurônio pós-sináptico tenha mais dificuldade em disparar potenciais de ação (LENT,
2001).
Como o neurônio pós-sináptico recebe inúmeras ações sinápticas, sua atividade
depende do somatório de todos os potenciais gerados por todas as sinapses que ocorrem
em sua membrana. Este processo de somação é denominado de integração sináptica.
Por exemplo, o potencial de repouso da membrana do soma neuronal de alguns tipos de
neurônios é de cerca de -65 mV (Figura 2.12). Na sinapse excitatória, este potencial
sobe para -45 mV (potencial pós-sináptico excitatório – PPSE). Entretanto, a descarga
de um só terminal não é capaz de elevar o potencial desta forma, assim, são necessárias
várias descargas simultâneas de muitos terminais, por exemplo de 40 a 80 terminais
para um neurônio motor comum (GUYTON, 2002). Na sinapse inibitória, o potencial

47
cai a -70 mV (potencial pós-sináptico inibitório – PPSI), impedindo a ação do neurônio
pós-sináptico, ou seja, impedindo que os outros neurônios possam ativá-lo.
Figura 2.12 – Eventos eletroquímicos da sinapse excitatória e sinapse
inibitória (adaptada de (GUYTON, 2002))
2.5 Medidas Neurofisiológicas: Matrizes Multieletrodo
(MEA)
2.5.1 Culturas celulares
Em condições apropriadas, a maior parte das células vegetais e animais poderão
viver, multiplicar-se e até mesmo expressar propriedades diferenciadas em uma placa de
cultura de tecidos. As células podem ser observadas sob o microscópio e várias análises
analisadas bioquimicamente, e os seus aspectos morfológicos, imunológicos e
farmacêuticos podem ser explorados. Experimentos com células oriundas de cultura são,
às vezes, ditos como tendo sido conduzidos in vitro para contrastá-los daqueles
experimentos com organismos intactos, os quais são referidos como conduzidos in vivo.
Os termos podem ser confusos porque são freqüentemente utilizados com sentidos
diferentes por bioquímicos, para quem in vitro é aplicado em referência a reações
48
bioquímicas ocorrendo em cultura; enquanto in vivo é aplicado para qualquer reação que
ocorra dentro de célula no seu ambiente natural.
A técnica de cultura celular foi inaugurada por Ross Granville Harrison, no
início do século XX (BEADLE, 1988). Ele realizou experimentos com sapos e
posteriormente com pintinhos, mostrando que fibras nervosas desenvolvem-se a partir
de corpos celulares individuais, ao contrário do que até então se acreditava. Entretanto,
há apenas trinta anos atrás, os métodos de cultura de células começaram a ganhar
proeminência na neurobiologia, motivados por duas descobertas. A primeira, os
neuroblastos, são células embrionárias do tecido nervoso que originam outras células
nervosas, obtidos de um tumor, que podiam ser investigados e controlados quando
mantidos em cultura, de forma a induzir a diferenciação neural (SCHUBERT, 1969). A
segunda, as culturas monitoradas in vitro, pela elongação axonal (BRAY, 1970).
Existem basicamente três métodos para cultura de células: o uso de células
dissociadas, cultura organotípica e de reagregação (BANKER, 1991). A cultura de
células dissociadas ou dispersas apresenta como vantagem a manutenção das
propriedades morfo e fisiológicas presentes no tecido de origem. A cultura organotípica
se refere à transplantação de tecido; sua vantagem principal é a preservação da
arquitetura do tecido. Por outro lado, esta técnica é aplicável apenas se o tecido é ou
pode ser manipulado na forma laminar. As células reagregadas são dissociadas e
mantidas em suspensão; elas tendem a se reagregar em pequenas esferas se mantidas em
frascos e centrifugadas. Este tipo de cultura é adequado para o estudo do
amadurecimento neural (STIER, 1993). A técnica mais adequada para avaliação de
crescimento axonal e formação de sinapses entre neurônios é a dissociação. Nesta
técnica, as culturas podem ser primárias, sendo utilizadas nos experimentos diretamente
logo após extração, sem qualquer alteração; e de linhagens celulares, em que o processo
de mitose é induzido. Neste último caso, as células se multiplicam originam várias
gerações, criando uma população com características genéticas homogêneas e bem
conhecidas.

49
2.5.2 Interface bioeletrônica
Uma interface bioeletrônica corresponde à um sistema onde um pedaço de tecido
nervoso, geralmente mantido vivo em cultura celular, é conectado a um circuito elétrico,
na tentativa de se estabelecer um intercâmbio bidirecional de informação.
A Figura 2.13 ilustra o funcionamento dessa interface, ou seja, a integração do
sistema neural à eletrônica digital (BONIFAZI, 2002). No primeiro passo (a), é ilustrada
a interface individual das células nervosas. No próximo passo, o par de células nervosas
é acoplado. No arranjo (b), a estimulação do neurônio A induz a transferência do
potencial de ação pela rede neural para o neurônio B, cujo sinal é gravado através do
transistor. No arranjo (c), o registro do sinal do neurônio A ocorre pelo transistor, sendo
transferido posteriormente para o chip de microeletrônica, até atingir o neurônio B, que
é então estimulado. No último passo (d), as redes neurais são definidas e a criação do
chip para a comunicação é estabelecida, fornecendo uma rede dinâmica e computacional
para as comunicações bidirecionais entre os sinais. Portanto, a conexão neurônio –
circuito – neurônio é o passo elementar da interface bioeletrônica.
(a)
(b)
estimulador
transistor
Neurônio
A
Neurônio
B
Neurônio
B
estimulador
transisto
r
Rede Neuronal
Neurônio
A
(c)
(d)
Dinâmica Neural
Neurônio
Neurônio
A
B
Sistema Digital
estimulado
r
Microeletrônic
a
transistor
Figura 2.13 – Interface bioeletrônica (adaptada de (FROMHERZ, 2003))
Dentro do que foi acima exposto e, considerando dois circuitos híbridos com
neurônios baseados em chip de silicone (arranjo (c) da Figura 2.13), a Figura 2.14
ilustra a dinâmica de cada neurônio A e B. No item (a) desta última figura, apresenta-se

50
o potencial de ação do neurônio A, correspondente à atividade elétrica basal ou
espontânea, a qual é captada e amplificada por um circuito, gerando a onda (c). Em
seguida, os picos da última onda, correspondentes à ocorrência de um potencial de ação
no neurônio A, são detectados através de algoritmos de processamento, resultando na
onda (e). Após um ajuste de amplitude e de freqüência, de forma a garantir uma
coerência biológica, o sinal (e) é transformada em (f) através de circuitos lógicos, os
quais, finalmente, geram uma onda de estimulação (d). Esta última é aplicada ao
neurônio B através de outra interface bioeletrônica, induzindo potenciais de ação em B,
como mostrado em (b).
Figura 2.14 – Caminho neurônio- chip - neurônio (adaptada de (FROMHERZ, 2003)).
Vide arranjo (c) da Figura 2.13
2.5.3 Matrizes Multieletrodo (MEA)
A atividade elétrica em sistemas biológicos é tradicionalmente monitorada com
eletrodos, como por exemplo, na técnica de “patch-clamp”(GUEDES, 1989).
Matrizes Multieletrodo correspondem a um refinamento do conceito de interface
bioeletrônica ((FROMHERZ, 2003), (BONIFAZI, 2002)), e consistem em circuitos de
dimensões micrométricas montados sobre um substrato, e dotados de um conjunto de
eletrodos, os quais são colocados no soma do neurônio (Figura 2.15). A condição ideal
para o uso destas estruturas exige que o corpo celular se localize sobre um eletrodo, ou
entre dois eletrodos vizinhos. Para que isto ocorra, é necessário o posicionamento da
célula no local do eletrodo, bem como o tratamento adequado do substrato, de forma a

51
promover a aderência celular, ou seja, o substrato é uma das variáveis a serem
consideradas no processo de fabricação.
Microeletrodo
Tecido
Nervoso
Substrato
Figura 2.15 - Conjunto de eletrodos em um tecido
nervoso (adaptada de (RENNAKER, 2005))
As MEAs são utilizados para acessar de forma sistemática e seletiva os
potenciais de ação associados a feixes ou fibras nervosas, sendo que vários testes em
modelos animais já são realizados em diversos centros de pesquisa (URL 5). Elas
permitem registrar simultaneamente a atividade elétrica de cerca de centenas de
neurônios. Os eletrodos da MEA podem também ser usados para estimular neurônios,
de forma não-invasiva, seguindo uma conexão entre a rede neuronal de cultura e um
computador.
A Figura 2.16 ilustra a aplicação de MEAs em outros métodos de cultura. Na
primeira (a), um extrato da massa branca do cerebelo é posicionado sobre um substrato
contendo microeletrodos, para extração dos sinais. Já na segunda (b), os eletrodos
(representados por hastes negras) são posicionados sobre os axônios de embriões de
ratos, após 14 dias em cultura, evidenciando o número de células acessíveis que foram
atingidas.

52
Eletrodos
Cerebelo
(a) (b)
Figura 2.16 - Exemplos de aplicação das matrizes de microeletrodos
(adaptada de PEIXOTO, 2001) (a) corte de cultura em tecido do cerebelo sobre
MEA, (b) cultura celular de neurônios isolados do gânglio da raiz dorsal de ratos em
cultura (14 dias).
2.5.4 Utilização e Fabricação das MEAs
Quando se mede a atividade nervosa de células individuais ou conjuntos bem
determinados de células, seja em cultura in vitro ou in vivo, utilizam-se eletrodos
micrométricos, os microeletrodos, que são fabricados em metal ou vidro, com pontas de
diâmetros variando de décimos a dezenas de micrômetros (PEIXOTO, 2001).
Existem duas formas de medida de microeletrodos: intra e extraceleular. Os
microeletrodos para medidas intracelulares são fabricados a partir de capilares de vidro
e preenchidos com solução salina de alta molaridade. Estes eletrodos assim preparados
são denominados de micropipetas e são projetados para inserção na membrana celular,
ou para formar um selo através de sucção mecânica na membrana. O princípio de
medida se baseia na capacitância da parede de vidro e na alta concentração iônica da
solução salina interna do eletrodo. Já para sinais extracelulares, eles são classicamente
adquiridos através de eletrodos de tungstênio, também denominados de Wolfram, e com
micropipetas.
O posicionamento do microeletrodo é o fator determinante para a obtenção de
um bom resultado na medida eletrofisiológica. Além disso, o tempo de medida é
limitado, impedindo, por exemplo, a utilização de circuitos híbridos bioeletrônicos por
mais do que algumas semanas.
As MEAs têm sido desenvolvidas e diversificadas, como por exemplo com a
incorporação de dispositivos eletrônicos integrados e a fabricação tridimensional de

53
eletrodos (Figura 2.17). Deve-se procurar manter os requisitos essenciais de
biocompatibilidade, estabilidade mecânica, manutenção do ambiente fisioquímico
(como a temperatura, o pH, a composição da solução fisiológica e a esterilidade)
durante o experimento. Logo, as principais vantagens destas matrizes são a fabricação
em larga escala de microeletrodos e a utilização de sistemas de aquisição que não estão
restritos ao posicionamento fino através de micromanipuladores.
Figura 2.17 – Matriz tridimensional fabricada com técnicas de microeletromecanismos
(adaptada de (BAY, 2000))
Uma matriz de microeletrodos pode ser implementada sobre um substrato,
usualmente de vidro ou silício (vide Figura 2.18), sobre o qual uma camada metálica é
construída por meio de técnicas fotolitográficas, sendo protegida por um material
isolante, através da camada passivadora. Nesta última, são abertas janelas através das
quais o metal fica exposto à cultura. A área exposta ao metal é denominada de
microeletrodo (PEIXOTO, 2001).
Figura 2.18 – Esquema geral da fabricação da MEA
(adaptada de (PEIXOTO, 2001))
54
A implementação de microeletrodos baseia-se em técnicas tradicionais de
microeletrônica, que são aplicadas com êxito relativo, devido à restrições de
encapsulamento e ao tempo de estabilidade reduzido. Estas limitações são justificadas
pela presença de soluções fisiológicas em contato com o circuito.
Os eletrodos devem ser biocompatíveis, duráveis, e apresentar uma impedância
baixa (abaixo de 500 Ω a 1 kHz), para conseguir captar pequenos sinais extracelulares,
cuja amplitude atinge valores de ordem de 10 a 100 microvolts. A baixa impedância
também permite injetar uma corrente estimulátoria, suficiente para atingir o tecido
nervoso sem exceder a baixa voltagem eletromecânica da água, ou de outros
componentes do meio. Os eletrodos têm sido recobertos por poros de platina, de baixa
durabilidade, aumentando assim a impedância, de forma inaceitável. Esse problema
pode ser reduzido pelo uso de recobrimento de platina do tipo “sonicating” (PEREIRA,
2004).
Em relação ao problema de assegurar baixa impedância, os eletrodos encapados
são fabricados à base de oxido de irídio ou de nitrito de titânio. A superfície da MEA e
as pontas dos eletrodos devem ser recobertas com algum protetor compatível, que
também deve promover a adesão celular. Este protetor leva em conta os substratos
tradicionais da cultura celular, como os poliaminoácidos e laminina. A fabricação de
MEAs sem silicone permitiu alguns sucesso na estimulação do crescimento neuronal,
principalmente através da inclusão de substâncias neurotróficas.
Apesar da aparente simplicidade na fabricação, a escolha de materiais, a
caracterização das estruturas, a adequação para cada aplicação e propostas de modelos
são temas correntes na literatura quando se trata de interfaces neurais para aplicações,
seja in vitro ou in vivo ((BOVE, 1997), (HAIDARLIU, 1995)). Na maioria destes
sistemas, o projeto de uma microestrutura é precedido de um estudo sobre a aplicação
desejada, de um modelo do comportamento esperado e de propostas para a análise de
dados associadas aos experimentos, principalmente quando se trata de questões
específicas em neurociências.
2.5.5 Histórico
55
Considera-se que a primeira MEA foi proposta em 1972 por Thomas et al., ou
por Ken Wise em 1975 (WEIS, 1996), mas apenas em 1980 obteve-se a confirmação
experimental de que as matrizes de fato medem sinais extracelulares, que são os
correspondentes temporais dos sinais intracelulares. Pine (1980), usando uma MEA
com passivação de óxido de silício, mostrou a validação do sistema através de medidas
simultâneas intra e extracelulares. Em 1984 (ISRAEL, 1984), obteve-se pela primeira
vez a estimulação e a aquisição simultâneas na mesma cultura, fato até então inédito
com o uso de MEAs. Também se realizou implante de retina em uma mulher cega
utilizando MEAs de 80 eletrodos (BRINDLEY, 1968). O implante não apresentou
efeito clínico significativo, mas os resultados foram positivos no aspecto de segurança a
longo prazo do circuito.
Os primeiros resultados experimentais relativos à interface bioeletrônica foram
divulgados em 1991 e 1995, considerando células nervosas de sanguessuga acopladas a
transistores abertos, bem como a estimulação através de chip de silicone baseados em
circuitos integrados ((FROMHERZ, 1995), (BONIFAZI, 2002)). Os autores
estabeleceram um sistema otimizado em interface de silicone neural. Outro ponto
relevante foi a utilização de sistemas híbridos baseados em redes neurais acoplados em
circuitos de microeletrônica (BONIFAZI, 2002), (FROMHERZ, 1997). Desta maneira,
a supervisão de neurônios em uma rede por contatos não-invasivos em um substrato
semicondutor possibilitou estudos do processo neurodinâmico, tanto para leitura como
para gravação dos dados. Baseado nestes resultados, surgiram os primeiros
neuroimplantes utilizando as técnicas de MEAs. Um exemplo desta aplicação é o
implante em nervo fibular, induzindo neuromodulação para o controle da dor
(RUTTEN, 1999).
Outros estudos baseiam-se em interfaces de redes neurais, como exemplo a
prótese neural (SHAHAF, 2001). Além disso, deve-se destacar que a excitação elétrica
do tecido neural foi usada para muitos propósitos. Como exemplos, podem-se citar o
marca-passo, o implante de cóclea (GRILL, 1999) e as próteses visuais (VERAART,
1998). Interfaces cérebro-máquina (PFURTSCHELLER, 2001) também estão sendo
explorados na medicina em termos de excitação elétrica, como terapia da doença de
Parkinson (MOGILNER, 2001), bem como para reabilitação e “biofeedback”.
56
A tecnologia MEA é acessível apenas por alguns laboratórios, que não podem
implementar circuitos de micro-fabricação eletrônica. O sistema completo de registro,
com 60 eletrodos, é produzido pelas empresas Miltichannel Systems (Alemanha);
Panasonic (Japão); Guenter Gross associada à Univ. of North Texas, Plexon Inc (USA).
Recentemente, foi patenteado um sistema de aquisição de dados, incluindo MEAs de 60
eletrodos, com freqüência de aquisição da ordem de 20 kHz (PEREIRA, 2004).
2.5.6 Aplicações
Devido à quantidade de sinais adquiridos com a aplicação de matrizes
multieletrodo, vários sistemas dedicados foram desenvolvidos nos últimos anos e são
comercializados em conjunto com pré-amplificadores, filtros, microscópios, etc
(PEIXOTO, 2001). No caso particular de neurônios em cultura, os pontos importantes a
serem considerados na aquisição e pré-classificação de sinais são as seguintes
características: freqüência de amostragem, o número de canais e filtros utilizados, a
relação sinal/ruído e o método de discriminação dos potenciais de ação ((FROMHERZ,
1994), (GROSS, 1993)).
Estas aplicações podem ser classificadas em:
(A1) Análise da codificação neural
Tendo em vista a disponibilidade comercial de MEAs, diversos centros de pesquisa
estão registrando continuamente o código neural, gerando uma quantidade explosiva de
dados elétricos ou ópticos que necessitam de análise em tempo real (PEREIRA, 2004).
Logo, esta tarefa exige frequentemente a utilização de vários processadores e
computadores ao mesmo tempo.
A velocidade dos computadores permite realizar a análise e a geração de
estímulos para a simulação do comportamento de um tecido nervoso num determinado
ambiente, bem como da interação com ele. Logo, através da seqüência de estímulos
sensoriais artificiais, a cultura neuronal reage segundo padrões de atividade elétrica, de
tal forma que a atividade da rede biológica sofre mudanças persistentes, as quais podem
se tornar compreensíveis para o manipulador externo. A alta resolução de uma imagem,
adquirida em um pequeno lapso de tempo, usando simultaneamente vídeo-microscópio
ou fóton, permite estudar correlações morfológicas deste aprendizado biológico. Ao

57
mesmo tempo, a utilização de culturas contendo desde poucas dezenas a centenas de
células, pode ser considerada simples, permitindo assim a indução de padrões de
aprendizado, de reconhecimento e de processamento.
Outros avanços estão sendo desenvolvidos utilizando MEAs planares
(KRAUSE, 2003), para criar ou melhorar métodos que avaliam as combinações em
cultura. Neste artigo, o autor desenvolveu três metodologias, que se baseiam nos
diferentes aspectos das MEAs. Na primeira, ele utiliza o processamento paralelo,
quando otimiza as questões de hardware, software e outras tecnologias de incubação,
para realizar medidas de várias culturas simultaneamente. Na segunda, existe uma
combinação de MEAs e aplicações em cultura para o estudo de substâncias
neurotóxicas. Na última, utiliza-se uma MEA planar bidimensional. Com espaçamento
adequado entre os eletrodos, pode-se estudar múltiplas regiões corticais
simultaneamente, considerando apenas uma combinação para a saída obtida. Como
exemplo, pode-se citar o estudo de neuroendocrinologia, que revela as desordens
ligadas à obesidade.
A Figura 2.19a exemplifica o tipo de sinal obtido com a matriz de
microeletrodos planares. Em outro exemplo (vide Figura 2.19b), observam-se tanto
sinais extracelulares medidos pela matriz quanto os respectivos sinais intracelulares
obtidos com eletrodos de vidro.
Figura 2.19- (a) Matriz de microeletrodos com neurônios e sinais extracelulares medidos de vários
neurônios em cada eletrodo. Gânglio abdominal de Aplysia (adaptada de (STENGER, 1994)). (b) Sinais
58
de uma matriz extracelular versus sinais medidos com eletrodos intracelulares (adaptada de (BEADLE,
1988))
Um outro método para análise da codificação neural utiliza uma matriz de
multieletrodo com três dimensões, composta por microchips com submatrizes de
diferentes configurações (HUAI-YUAN, 2005). Essa inovação é desenvolvida para
aplicações médicas. O processo de fabricação três-máscaras é baseado em “wafer”,
utilizando os eletrodos em linhas e blocos. Os eletrodos são cobertos com SiO
2
de 2μm
de espessura para reduzir o ruído por oxidação, e foram testados em células nervosas da
retina do coelho.
(EGERT, 1996) desenvolveu suas pesquisas utilizando MEA para a análise do
potencial de ação de eletroretinograma (ERG). Ele concluiu que as características de
correlação entre os sinais ERG e o respectivo pico são elevadas, revelando alterações do
estado fisiológico do tecido. O autor também realizou testes com MEAs planares,
concluindo que elas são apropriadas para a gravação do sinal de pico e para potenciais
de baixa freqüência.
(A2) Neuroimplantes
Na Figura 2.20 pode-se observar um exemplo de aplicação terapêutica da MEA,
que é implantada em um nervo fibular, com o objetivo de realizar eletroestimulação
para induzir neuromodulação. Como o funcionamento dos eletrodos é flexível, eles
podem ser tanto usados para conduzir um impulso elétrico para dentro do nervo, quanto
para captar a atividade elétrica do nervo.

59
Figura 2.20 – MEA funcionando como neuroimplante em um nervo fibular (adaptada de (RUTTEN,
1999))
Implantes clássicos, como o marca-passo, caracterizam-se por um
funcionamento fixo, pré-determinado, estabelecido antes da intervenção cirúrgica de
implantação. Na literatura (BERGER, 2001), já existe a proposição de um
neuroimplante “inteligente”, adaptável às condições especificas do paciente. A Figura
2.21 apresenta um esquema simplificado deste neuroimplante. Uma vez os potenciais de
ação adquiridos e corretamente traduzidos pela interface bioeletrônica, é necessário
amplificá-los e formatá-los, para posteriormente realizar processamento de sinais. Este
último objetiva não apenas modelar fenômenos, mas também atuar na informação
biológica, com objetivos terapêuticos. O resultado deste processamento deve ser
formatado “biologicamente”, em termos das amplitudes de tensão e de freqüências, para
que, através de uma interface bioeletrônica, retorne ao mundo biológico.
Interface
Bioeletrônica
Amplificação e
Formatação
Processamento
de sinais
Conversor
D/A
Formatação
Biológica
Interface
Bioeletrônica
Interface
Bioeletrônica
Fibra
nervosa
Figura 2.21 – Esquema de um neuroimplante inteligente (adaptada de (PEREIRA, 2004))
60
Deve-se enfatizar que os neuroimplantes foram desenvolvidos para prevenção de
crises epilépticas, m
icos coerentes com a variabilidade
9 patíveis, o que se traduz, dentre outros
um microchip de silicone baseado em
3) Farmacologia
das aplicações farmacológicas, existem diversas características
portantes que fazem da MEA um
rônios por um longo período de
9 entação;
anais.
eficiente, reduzindo o número
de experimentos nos animais.
inimizando-as de acordo com experimentos preliminares de eletro-
estimulação realizados em animais (LITT, 2003). Neste contexto, deve-se destacar que
os dois desafios tecnológicos mais importantes para a concretização do uso clínico de
neuroimplantes consistem em (Proc. IEEE, 2001):
9 O desenvolvimento de modelos matemát
biológica do organismo humano;
A implementação de dispositivos biocom
quesitos, por mínima dissipação de potência. De fato, a amplitude desta em
microprocessadores comuns excede em diversas ordens de grandeza os níveis de
energia do encéfalo, levando à possibilidade de grave lesão do córtex caso
circuitos comuns sejam implantados.
O autor (SONG, 2005) desenvolveu
MEAs, com tecnologias CMOS, projetado para substituir uma região específica do
cérebro. O circuito integrado constitui-se de dezesseis pré-amplificadores de baixo
ruído, com multiplexadores e um amplificador na saída, para extração dos sinais neurais
durante a gravação. O neuroimplante foi testado em cérebros de ratos, avaliando-se as
medidas do potencial de ação.
(A
No contexto
im a ferramenta de medida eletrofisiológica relevante
(CHIAPPALONE, 2003), dentre as quais se destacam:
9 O monitoramento da atividade elétrica dos neu
tempo;
A forma não-invasiva de instrum
9 A gravação simultânea de até dezenas de c
Deste modo, MEAs permitem um processamento
61
A tecnologia MEA pode ser aplicada em tecidos eletrofisiológicos, por exemplo,
neurônio central, células do coração e células musculares. O biossensor MEA constitui
um sis
rface cérebro-máquina
Os sistemas de interface cérebro-máquina baseados nos sinais
estigados há 20 anos atrás (ROUSCHE, 2003).
alm
Doenças neurodegenerativas, como a esclerose múltipla, podem induzir a perda
. Desta forma, a implantação de biossensores
sis
tema ideal in vitro para monitorar os efeitos crônicos e agudos de drogas e
toxinas, e apresenta estudos funcionais induzindo condições patológicas sem danos. A
gravação da resposta elétrica é realizada em vários locais de um tecido, e portanto um
mapa de diferentes drogas pode ser gerado, provendo conclusões importantes sobre a
atuação bioquímica específica da droga em estudo (STETT, 2003). Este último artigo
apresenta uma revisão que descreve pesquisas com aplicações de biossensores MEA
para blindagem de drogas, bem como a segurança da farmacologia nos casos cardíaco e
neural.
(A4) Inte
eletroencefalografia foram inv
Atu ente, poucos sistemas são desenvolvidos de forma robusta para atender às
funcionalidades clínicas do dia-a-dia (WOLPAW, 2002), por exemplo, em ratos
(CHAPIN, 1999). ROUSCHE (2003) introduziu um sistema para gravações em ratos,
implantando-se MEAS em seu córtex, gerando um “robô de rato”, o RABOT. Ele é um
sistema especial para experimentos em neurociência e engenharia neural, visto que
induz a locomoção de pequenos robôs a partir da aquisição de sinais do córtex motor de
ratos. Vários testes movendo o RABOT por um som fixo levam a interessante respostas
neurais, possibilitando um grande avanço em aplicações cérebro-máquina. Futuros
experimentos com este sistema incorporam mecanismos de decisão complexa.
(A5) Implante medular e nanotecnologia
da funcionalidade no organismo humano
em temas neurais para a eletro-estimulação tem sido aumentada a cada dia. A
neurotecnologia utiliza dispositivos protéticos para suplemento ou substituição de
funções do sistema nervoso ((PEYMAN, 1998), (ZRENNER, 1999)).
62
Um exemplo desta abordagem é apresentado por (SCHLOSSHAUER, 2001),
que desenvolveu implantes neurais com eletrodos pequenos para a excitação externa de
unção de partes coordenadas para a
maneira a formar um conjunto inter-
as e internas e
produzir respostas a essas variações através dos músculos e glândulas. Desta
No sistema nerv
neuroglia (células de que desempenham a
neurônios co-implantados, cujos axônios penetram nos eletrodos e ativam as fibras
nervosas.
2.6 O Sistema Nervoso (SN)
Um sistema se caracteriza pela j
concretização de um mesmo resultado, de
relacionado. O Sistema Nervoso (SN) é, pois, um conjunto de órgãos formados por um
mesmo tipo de tecido, engajados num mesmo processo (GUYTON, 2002). Ao lado do
Sistema Endócrino, constituído por glândulas secretoras de hormônios, o Sistema
Nervoso é responsável por todas as funções de controle do corpo humano.
“O sistema nervoso humano é o mais complexo entre os animais. Sua função
básica é de receber informações sobre as variações extern
forma ele contribui, juntamente com o sistema endócrino, para a homeostase
do organismo. Além do mais, o sistema nervoso humano possui as chamadas
funções superiores que inclui: a memória, que corresponde à capacidade de
armazenar informações e depois resgatá-las, o aprendizado, o intelecto, o
pensamento e a personalidade” (VILELA, 2005).
oso, além dos neurônios que o constituem, têm-se as células de
suporte, também denominados gliócitos),
função de sustentar, proteger, isolar e nutrir os neurônios (VILELA, 2005). Essas
células são muito mais numerosas que os próprios neurônios, entretanto seus corpos
celulares são geralmente menores (LENT, 2001). Há diversos tipos celulares, mas se
distinguem, entre elas, os astrócitos, oligodentrócitos e microgliócitos (Figura 2.22).

63
Figura 2.22 – Células gliares: astrócitos (nutrição), oligodentrócitos
(sustentação) e células microgliais (defesa) (adaptada de (VILELA, 2005))
O SN se divide em Sistema Nervoso Central (SNC), Sistema Nervoso Periférico
(SNP) e Sistema Nervoso Autônomo (SNA), conforme a Figura 2.23. O Sistema
Nervoso Central pode ser subdividido em:
9 encéfalo, que corresponde à parte que está dentro da caixa craniana, e
9 medula espinhal, que corresponde à parte protegida pela coluna vertebral.
O Sistema Nervoso Periférico pode ser subdividido em:
9 nervos raquidianos, corresponde os nervos que partem da, ou chegam na,
medula,
9 nervos cranianos, corresponde aos que parte da, ou chegam na, região craniana,
9 Sistema Nervoso Autônomo, que é responsável pelo controle neurovegetativo –
desprovido do controle da nossa vontade.

64
Figura 2.23 – Figura esquemática mostrando o SNC, representado pelo
encéfalo e pela medula e o SNP, representado pelos nervos (adaptada de
(FRISÓN, 1999))
O SNC é responsável pela tomada de decisões. Suas funções são de receber,
avaliar e processar os estímulos sensitivos; como também de ponderar e enviar os
estímulos motores. O SNP é responsável pelo “sentir” o mundo externo e o mundo
interno (ao corpo), enviando estas informações ao SNC; como também se
responsabiliza por levar as informações do SNC para músculos e glândulas, que
efetuam suas tarefas de acordo com a tomada de decisão.
2.6.1 Sistema Nervoso Central (SNC)
O SNC é envolvido por estruturas ósseas protetoras, como a caixa craniana, que
protege o encéfalo; e a coluna vertebral, que contém a medula (Medula Espinhal ou
Medula Nervosa ou Ráquis). Além disso, o SNC está imerso numa substância aquosa
denominada líquor ou líquido cefalorraquidiano, e revestido por membranas
denominadas meninges, que são divididas em três (vide Figura 2.24): dura-máter (a
mais externa), aracnóide (a intermediária) e pia-máter (a mais interna).

65
Figura 2.24 – Meninges: Dura-máter (mais externa), Aracnóide
(Intermediária) e Pia-máter (mais interna)(adaptada de (CARDOSO, 2005))
Todo este líquor que banha o sistema nervoso central, preenchendo inclusive as
cavidades encefálicas (ventrículos), “o protege mecanicamente contra traumatismos
que possam atingir a cabeça, e também contribui com a sua nutrição e a manutenção
do meio bioquímico ótimo para o funcionamento neural” (LENT, 2001).
A dura-máter é extremamente resistente, rígida e bastante sensível. A aracnóide
é menos rígida que a dura-máter, entre as duas há uma fina camada de líquor que
lubrifica o contato entre ambas. Entre a aracnóide e a pia-máter, existe um amplo espaço
preenchido pelo mesmo líquor, o espaço subaracnóideo. A pia-máter, última das
meninges, é a mais fina e delicada, cobrindo todo o SNC, acompanhando, inclusive,
seus giros e sulcos (LENT, 2001).
2.6.1.1 A medula espinhal
A medula tem cerca de 40 cm de comprimento (vide Figura 2.25) em um
indivíduo adulto mediano. Ela é condutora dos impulsos nervosos que provêm do
encéfalo ou vão para este. Dela partem as raízes motoras e sensitivas dos nervos
raquidianos, levando informações para órgãos efetores e trazendo informações sobre o
que acontece com o corpo.

66
Figura 2.25 – Medula Espinhal e Encéfalo (adaptada de (CARDOSO, 2005))
A medula também funciona como uma espécie de “hardware” de controle,
atuando nos processos para os quais o sistema de processamento encefálico não precisa
agir de imediato. São funções simples, e portanto não necessitam da intervenção dos
centros encefálicos. Assim, a medula controla os movimentos involuntários como o
arco reflexo. Podemos citar, por exemplo, o ato de retirar a mão involuntariamente de
um objeto que tenha causado dor. A retirada da mão é imediata, mas a tomada de
consciência do que ocorreu vem após. O sinal de dor é transmitido a regiões especificas
da medula que processam a informação e a envia aos grupos musculares responsáveis
pela flexão do braço.
“Na realidade, os níveis superiores do sistema nervoso operam
freqüentemente, não mandando sinais diretamente para a periferia do corpo,
mas enviando sinais para os centros de controle da medula, simplesmente
“comandando” os centros da medula espinhal a desempenhar suas funções”
(GUYTON, 2002).

67
2.6.1.2 O encéfalo
Todo o material composto pelo tecido nervoso situado dentro da caixa craniana
constitui o encéfalo (Figura 2.26). Ele é formado pelo:
9 Nível encefálico inferior, responsável pelo controle de atividades orgânicas
inconscientes, são eles:
(a) diencéfalo – tálamo, hipotálamo, epitálamo e subtálamo
(b) tronco cerebral - mesencéfalo, ponte e bulbo
(c) cerebelo.
9 Nível encefálico superior, responsável pela coordenação de atividades
conscientes, relacionadas às atividades intelectuais e voluntárias, que são:
(a) telencéfalo – os hemisférios cerebrais.
Figura 2.26 – O encéfalo (adaptada de (URL 6))
Diencéfalo
No diencéfalo encontramos o tálamo, constituído de duas massas ovóides de
substância cinzenta e branca (Figura 2.27), e o hipotálamo. Essas estruturas são
responsáveis por funções como controle do medo, do sono, da digestão, da fome, da dor
e da regulação térmica dentre outras funções de controle da nossa homeostase.
O tálamo faz parte do sistema límbico (hipotálamo, tálamo, amígdala,
hipocampo, corpos mamilares, cíngulo, giro parahipocampal), que regula o
comportamento emocional, memória e aprendizado. Com exceção das mensagens

68
sensoriais provenientes do bulbo olfativo, todas as outras passam diretamente pelo
tálamo antes de atingir o córtex cerebral. O tálamo funciona como um filtro, tendo a
função de integração sensorial motora, e retransmite os impulsos nervosos para o córtex,
direcionando-os às regiões especificas do cérebro onde devem ser processados.
Já o hipotálamo é o grande coordenador da homeostasia (LENT, 2001), definida
como um estado do organismo associado a:
9 ambiente interno contendo concentrações otimizadas de gases, nutrientes, água e
íons;
9 temperatura regulada;
9 volume de fluídos otimizado para a vida das células.
Ou seja, o hipotálamo busca manter o sistema em um nível estável, regulando-o
em resposta a flutuações aleatórias do ambiente, que não podem ser antecipadas.
Tronco encefálico
O mesencéfalo e a ponte (Figura 2.27) são vias de passagem dos estímulos nervosos,
principalmente aqueles relacionados à motricidade e à sensibilidade.
Figura 2.27 – Diencéfalo e Tronco Encefálico. Vista posterior do encéfalo
com exclusão do cerebelo (adaptada de (VILELA, 2005))
O Bulbo (Figura 2.28) também é via de passagem dos estímulos que vem do, ou
vão para o encéfalo, com funções próprias, como controle do espirro, da tosse, da

69
sucção, da fonação, da mastigação, da respiração e da salivação; além de possuir função
cárdio-moderadora, cárdio-aceleradora, vasoconstrictora e vasodilatadora. Deve-se
destacar os núcleos da formação reticular que acumulam as funções acima citadas.
Figura 2.28 – Ponte e Bulbo em evidência, com o cerebelo ao fundo e parte
do telencéfalo acima. Vista anterior do encéfalo (adaptada de CARDOSO,
2005))
Cerebelo
O cerebelo fica logo atrás da ponte e do bulbo (Figura 2.29) e, assim como o
cérebro, está recoberto por uma fina camada de substância cinzenta denominada córtex
cerebelar, cobrindo seu corpo branco medular.
Figura 2.29 – Secção mostrando a metade direita do encéfalo vista por
dentro, evidenciando suas subdivisões (adaptada de (VILELA, 2005))
70
“As principais funções do cerebelo são: manutenção do equilíbrio e da postura,
controle do tônus muscular, controle dos movimentos voluntários e aprendizagem
motora” (MACHADO, 2003); regulando assim as funções musculares, a orientação
espacial e o equilíbrio do corpo. De fato, o equilíbrio corporal, “é a função combinada
das porções mais antigas do cerebelo e da substância reticular do bulbo, da ponte e do
mesencéfalo” (GUYTON, 2002). Já para os movimentos voluntários, o cerebelo
controla a força, a extensão e a direção do movimento, atuando no planejamento e
correção dos movimentos.
2.6.2 Sistema nervoso Periférico (SNP)
O SNP é formado por estruturas fibrosas denominadas de nervos, que são
constituídos por vários feixes de fibras nervosas, as quais, por sua vez, são a reunião de
prolongamentos de neurônios (axônios ou dendritos). Os nervos constituem os
componentes principais do SNP e se encontram em quase todas as partes do corpo
humano (LENT, 2001).
O SNP é composto de 12 pares de nervos cranianos, 31 pares de nervos
raquidianos e pelo Sistema Nervoso Autônomo. Estes nervos são ditos cranianos ou
raquidianos em função do seu ponto de ligação com o SNC, ou seja, podem emergir da
base da caixa craniana ou do tronco encefálico, ou da medula espinhal, respectivamente.
2.6.2.1 Sistema nervoso Autônomo
Ele é responsável pelo controle involuntário da vida vegetativa, coordenando
funções como a circulação, a respiração e a digestão, podendo ainda ser subdividido em
Sistema Nervoso Simpático e Sistema Nervoso Parassimpático (vide Figura 2.30). Estes
atuam antagonicamente um em relação ao outro, porém de forma equilibrada. Por
exemplo, o Sistema Nervoso Simpático excita os órgãos efetores, enquanto o Sistema
Nervoso Parassimpático os inibe.

71
Figura 2.30 – Sistema Nervoso Central e Periférico. Evidência de parte do
Sistema Nervoso Autônomo (Simpático e Parassimpático), bem como de
exemplo de nervos motores e sensoriais (adaptada de (CARDOSO, 2005)).
2.6.3 Cérebro
O cérebro é dividido em dois hemisférios (esquerdo e direito), sendo que, com
relação às funções motora e sensitiva, um hemisfério coordena o lado do corpo que lhe é
oposto. Ou seja, o hemisfério cerebral direito se relaciona com o controle motor e
sensitivo do lado corporal esquerdo enquanto o hemisfério esquerdo se relaciona com
controle do lado direito do corpo. Assim, há funções cognitivas peculiares de cada
hemisfério, ambos se comunicando por meio de uma estrutura fibrosa denominada
corpo caloso, o qual se caracteriza pela substância esbranquiçada situada abaixo do
telencéfalo, na Figura 2.29. O corpo caloso cruza a fissura cerebral longitudinal, fenda
que divide o cérebro em dois hemisférios.

72
2.6.3.1 O córtex cerebral
Os hemisférios cerebrais apresentam uma camada superficial de 2 a 5 mm de
espessura de substância cinzenta, composta por neurônios, células de glia e fibras
nervosas, denominada córtex cerebral ou córtice cerebral.
“É irônico que, de todas as partes do cérebro, saibamos menos acerca das
funções do córtex cerebral, mesmo que esta componha, de longe, a maior
porção do sistema nervoso. Mas sabemos quais os efeitos da lesão ou da
estimulação específica em várias porções do córtex” (GUYTON, 2002).
Fisicamente, o córtex cerebral pode ser dividido em quatro lobos externamente
visíveis, que recebem o nome da estrutura óssea que os protege (Figura 2.31). O quinto
lobo é denominado de ínsula, localizando-se na parte interna e central do cérebro,
abaixo do lobo temporal.
Figura 2.31 – Lobos do córtex cerebral (adaptada de (VILELA, 2005))
Em geral, o lombo temporal se dedica à memória, audição, fala e sua
interpretação; o lobo frontal é responsável pelo planejamento consciente, controle motor
e controle sensorial somático; o lobo parietal lida com os sentidos corporal e espacial
do lado direito; o lobo occipital é uma área interpretativa do sentido da visão; e a ínsula
cuida da representação do estado fisiológico do corpo, envolvendo, inclusive, as
emoções. Portanto, é preciso esclarecer que os lobos cerebrais são interdependentes
através de circuitaria própria que os interconecta.

73
Existe uma relação bastante estreita entre o córtex e o tálamo, como se
existissem no tálamo as mesmas regiões funcionais do córtex. Todas as mensagens
sensoriais, com exceção daquelas provenientes do bulbo olfativo e motora, passam pelo
tálamo antes de atingir o córtex, e após sair do córtex cerebral. Impulsos oriundos das
vias da sensibilidade atingem o córtex, sendo interpretadas. Também do córtex partem
os impulsos que comandam os movimentos voluntários.
Todo o córtex está interconectado e suas regiões são interdependentes. As fibras
que penetram o córtex cerebral passam, necessariamente, pelo centro branco-medular
do cérebro, que constitui a substância branca revestida pelo córtex, composta por fibras
mielinizadas. Essas fibras podem ser de associação ou projeção. As fibras de
associação ligam áreas diferentes no mesmo hemisfério, ou no hemisfério situado no
lado oposto; enquanto aquelas de projeção ligam áreas do córtex aos centros
subcorticais, constituindo a cápsula interna.
Diante desta complexa interconexão do córtex, ele não constitui de uma
informação homogênea, sendo possível dividi-lo em diferentes áreas, de acordo com
suas funções, conforme a Figura 2.32.
Figura 2.32 – Subdivisões funcionais corticais, cerebelo e tronco encefálico
(adaptada de (VILELA, 2005))
74
A área da aquisição das palavras é a área de Broca, no giro temporal inferior
esquerdo, responsável pela fala. A área de compreensão das palavras é a área de
Wernicke, numa fronteira entre o lobo occipital temporal e parietal esquerdo. A área de
audição é uma área de audição primária no giro temporal anterior, uma vez que a
interpretação é feita na área de Wernicke. Todas as informações sensoriais que
necessitam da inteligência interpretativa são direcionadas das áreas sensórias para a área
de Wernicke. A região de comando motor, ou área motora voluntária, possui conexões
com os músculos efetores específicos, passando, como visto anteriormente, pelo tálamo
e em seguida por centros específicos da medula espinhal. Assim também, a região de
tato e sensibilidade, ou área sensorial somática, também possui conexões com células
sensoriais distribuídas pelo corpo, igualmente possuindo estações em centros
específicos da medula e do tálamo.
De acordo com MACHADO (2003), “o conceito das localizações funcionais do
córtex sofreu um abalo quando se consegui causar movimento por estimulação de áreas
corticais, tidas como exclusivamente sensitivas”. Logo, essas regiões foram mapeadas
de acordo com a especialização maior dessas áreas, mas não necessariamente são essas
suas únicas funções.
No entanto, o mapeamento dessas regiões segundo suas funções é usualmente
aceita, o que é bastante importante tanto para a compreensão do funcionamento do
cérebro, como para o diagnóstico de diversas lesões que podem ocorrer neste órgão.
2.7 Medidas Neurológicas: O Eletroencefalograma (EEG)
2.7.1 Introdução
As atividades neuronais do córtex cerebral ecoam sob forma de onda
eletroquímica até a superfície do escalpo, onde podem ser captadas através do EEG de
rotina, o qual dura 20-40 minutos e usa aproximadamente 20 eletrodos diferentes de
escalpo colocados de acordo com um determinado protocolo. Qualquer mudança
elétrica registrada na superfície cortical é causada pela atividade ordenada e simétrica de
uma classe de células do córtex cerebral. Além disso, um número suficiente de
75
neurônios deve disparar sincronicamente para gerar potenciais corticais (BUTTON,
2000).
A teoria mais aceita atualmente sobre a origem do sinal eletroencefalográfico,
proposta na década de 30, supõe que esses biopotenciais surgem do centro para o topo,
de dentro para fora do cérebro. Baseia-se na idéia de que o tálamo no diencéfalo vibra
continuamente e é esta vibração que faz com que o cérebro vibre ciclicamente (URL 7).
Trata-se de vibração no sentido de oscilação do potencial eletro-químico e não no
sentido mecânico.
Uma outra teoria, na qual os biopotenciais surgem do topo para o centro, de fora
para dentro do cérebro, é proposta pelo físico Paul Nunez (1995) (NUNEZ, 1995). Ela é
baseada na idéia de que o neocórtex, situada no topo dos hemisférios cerebrais, regula
as oscilações do córtex, que por sua vez faz com que o tálamo simplesmente ajuste a sua
própria oscilação à oscilação do neocórtex.
Nunez fundamenta esta idéia em investigações feitas por Steriade (STERIADE,
1993), que, em 1976, descobriu que quando o córtex motor era estimulado eletricamente
com períodos pequenos de 10 Hz a cada dois segundos, o tálamo começava a oscilar
espontaneamente com a mesma freqüência, ao fim de 28 períodos de estimulação.
Portanto, Nunez concluiu que a associação da atividade neocortical e aquela do tálamo
em mudar a sua freqüência fazem com que o tálamo e o cérebro harmonizem as suas
oscilações, que depois são capturadas pelo EEG (URL 7).
Portanto, a origem do EEG ainda é obscura. No entanto, sabe-se que possibilita
identificar uma série de patologias neurológicas e psiquiátricas, principalmente para
auxiliar no diagnóstico de doenças do cérebro, tais como a epilepsia, as desordens do
sono e alguns tipos de tumores cerebrais.
Fisiologicamente sabe-se que as características das ondas elétricas cerebrais
variam conforme o funcionamento do órgão. As maiores variações se observam entre os
estados de vigilância, ou seja, entre o estar acordado, dormindo, sonolento, em coma,
etc.
Uma dessas novas aplicações do EEG é tentar localizar com exatidão os focos
epilépticos ou tumores cerebrais. Os focos epilépticos são pequenas regiões no cérebro
onde a atividade elétrica se apresenta anormal. Pela observação dos traçados dos canais,

76
o neurologista que interpreta o EEG é capaz de deduzir em onde exatamente esta
anormalidade está situada. Logo, o EEG pode ser considerado o melhor exame
complementar para clínica, diagnóstico definitivo e terapia da epilepsia (SILVA, 1998).
2.7.2 Medição
Os sinais cerebrais são geralmente captados por meio de eletrodos em forma de
disco, devidamente posicionados no couro cabeludo (Figura 2.33), o que caracteriza
uma captação não-invasiva. Existem outras formas de captação invasiva dos sinais
cerebrais, por meio de agulhas subcutâneas, intracranianas, ou ainda redes de eletrodos,
que podem ser posicionados sobre o próprio córtex. Este procedimento é usado em
alguns casos de cirurgia neurológica.
Figura 2.33 – Eletrodos posicionados no escalpo e o registro
eletroencefalográfico (adaptada de (LENT, 2001))
É necessária uma substância ou suporte material que possa servir como
superfície fixadora entre o eletrodo e a pele do paciente, aumentando a aderência, de
modo que o eletrodo permaneça onde foi colocado. Isto permite que toda a superfície do
eletrodo possa captar o sinal, além de se evitar capacitâncias e resistências indesejáveis
associadas à movimentação desproposital do eletrodo. O paciente deverá estar sem
brincos ou objetos de metal que possam causar interferência, com os cabelos secos e o
77
couro cabeludo limpo. Cabelos longos devem ser presos de forma a exibir o escalpo. O
ambiente de registro deve ser calmo, silencioso, sem luzes intensas, sem movimentos de
pessoas, e obviamente, sem interferência eletromagnética. A aparelhagem deve ser
mantida muito limpa e freqüentemente calibrada.

77
Existem diversos tipos de eletrodos. Segundo a classificação de (BUTTON, 2000),
são eles:
• Eletrodos de disco de prata: eletrodos de superfície, possuindo um orifício para
a aplicação do gel condutivo. Possuem de 6 a 10 mm de diâmetro (Figura 2.34);
Figura 2.34– Eletrodos de disco de prata (modelos da Grass®) (adaptada de
(BUTTON, 2000))
• Eletrodos de agulha de platina: devem ser esterilizados e são feitos para
aplicação única, ou seja, são descartáveis. O comprimento da agulha de platina é
de cerca de 10 mm e pode ser longitudinal ou em ângulo reto. Estes eletrodos
são inseridos tangencialmente por debaixo da pele, apresentando o risco de
quebra e sangramento (Figura 2.35);
Figura 2.35 – Eletrodos de agulha de platina (modelos da Grass®)
(adaptada de (BUTTON, 2000))
• Eletrodos especiais:
o Nasofaríngeo: trata-se de um bastonete de prata, fino e flexível, de 10 cm
de comprimento para crianças e 15 cm para adultos, com a extremidade

78
captadora de ouro ou prata, de 3 mm de diâmetro. Deve ser inserido na
cavidade nasal em direção ao bulbo olfativo (Figura 2.36);
Figura 2.36 – Eletrodos nasofaríngeo (adaptada de (BUTTON, 2000))
o Grade de eletrodos: os eletrodos são prefixados num molde, que pode ser
uma grade ou uma touca (Figura 2.37). Podem ser do tipo ventosa de
borracha, através da qual se faz a fixação sem colódio, como para alguns
eletrodos mais antigos utilizados em eletrocardiograma; com um bastão
de Ag-AgCl (prata-cloreto de prata) no meio, ou possuir pequenos
orifícios para aplicação do gel condutor;
Figura 2.37 – Eletrodos prefixados em grade (adaptada de (BUTTON,
2000)). Touca com eletrodos posicionados (adaptada de (CARDOSO, 2005))
o Fios de prata ou aço inoxidável: são inseridos por uma agulha e são
usados em caso de implante de eletrodos por tempo prolongado, ou
durante monitoração de cirurgia. Os fios eletrodos são envernizados a
partir da ponta em forma de gancho. Quando o local de acesso é atingido,
a agulha é retirada deixando o eletrodo que fica preso pelo gancho;

79
o Clipe de orelha: é formado por dois eletrodos tipo disco (ouro ou prata),
com orifício para inserção do gel condutor, montados num suporte
plástico. São presos no lobo auricular e servem como eletrodo de
referência (Figura 2.38).
Figura 2.38 – Eletrodos tipo clipe de orelha (“ear clip”) (adaptada de
(BUTTON, 2000))
o Eletrodos Corticais: são fios flexíveis de ouro ou prata usados para
registrar potenciais elétricos no córtex exposto (Figura 2.39).
Figura 2.39 – Eletrodos corticais (adaptada de (BUTTON, 2000))
O sinal detectado pelo eletrodo deve ser devidamente condicionado para
posterior processamento. Por se tratar de um sinal de amplitude muito pequena, e por se
originar de um tecido vivo, o amplificador EEG precisa possuir alta impedância de
80
entrada e ótima razão de rejeição em modo comum. Os amplificadores mais comuns
utilizados nos equipamentos de EEG são de instrumentação, com impedância de entrada
da ordem de 10
12
Ω e razão de rejeição em modo comum (CMRR) maior que 500:1 (80
dB). Como a diferença de potencial medida por um par de eletrodos é de, tipicamente,
alguns poucos microvolts, existe a necessidade de um alto valor de amplificação, da
ordem de 20 000 a 100 000 vezes.
O número de canais que podem ser medidos simultaneamente varia de
equipamento para equipamento. Um eletroencefalógrafo pode ter 8, 16, até 128 ou 256
canais. O sinal amplificado deve ainda ser filtrado, enquadrando-se dentro da faixa de
freqüência de maior contribuição do sinal eletroencefalográfico registrado no escalpo
(0.5 a 30 Hz), separando componentes de freqüência indesejados. Em equipamentos
modernos, o sinal é então digitalizado e transmitido a um computador que o processa
e/ou armazena. Também é necessário um isolamento galvânico entre o paciente e a rede
elétrica, o que é geralmente realizado antes da filtragem do sinal.
2.7.2 Características estatísticas do sinal EEG
O sinal EEG é um sinal estocástico, ou seja, caracteriza-se por uma variação
aleatória dos valores de potencial elétrico no tempo.
As ondas cerebrais provêm dos potenciais de membrana resultantes de ações
sinápticas em células piramidais, que apresentam a disposição dipolar paralela ideal
para que seus potenciais sejam somados e registrados no escalpo. São ainda o resultado
do somatório da ação de milhares ou milhões de neurônios disparando sincronicamente
(CARDOSO, 2005).
Quando os neurônios disparam de forma assíncrona, os potenciais podem se
anular ao serem somados. Por este motivo, a amplitude do sinal EEG não diz respeito ao
nível total de atividade cerebral, mas principalmente à quantidade de neurônios
disparando sincronicamente. Já a freqüência da onda cerebral está diretamente
relacionada à atividade dos neurônios: quanto maior esta última, maior será a freqüência
de onda registrada (GUYTON, 2002).

81
A amplitude do sinal EEG pode variar de 0 a 200 µV, e a freqüência varia de um
pouco acima de zero a 30 Hz, salvo alguns estados patológicos em que esta freqüência
pode ser mais alta, como no caso de crises de epilepsia. Como a potência do sinal EEG
depende da quantidade e sincronicidade dos disparos neuronais, tais sinais são
considerados a medida da atividade neural de um volume relativamente grande de
muitos neurônios.
2.7.3 Tipos de ondas EEG
O espectro do sinal EEG pode ser subdivido em diferentes tipos de ondas: alfa,
(α), beta (β), delta (δ), teta (θ), gama (γ), lambda (λ), mi (µ), Ondas Fusiformes de Sono
e Complexos K. Observar na tabela 2.1 e na Figura 2.40 a descrição detalhada destas
ondas.
Tabela 2.1 – Ondas cerebrais (adaptada de (BUTTON, 2000))
Ritmo (onda) Freqüência A que se relacionam
Região cortical
mais comum
(em um adulto)
Delta
0,5 a 3,5
Hz
Sono profundo. Frontal
Complexos K
0,5 a 3,5
Hz
Ondas delta de amplitude
muito alta, com ápice
pontiagudo: sono leve.
Frontal
Teta 4 a 7 Hz
Hipnose; desenvolvimento da
memória; meditação;
sensações de estar fora do
corpo; experiências emocio-
nais; estresse; sonolência;
sonhos
Temporal e
parietal
Mi 7 a 11 Hz
Está numa intersecção entre a
faixa téta e a alfa: intenção de
mover-se.
Central e
temporal
(Córtex motor)
Alfa 8 a 13 Hz
Vigília com olhos fechados;
relaxamento mental;
meditação.
Occipital (às
vezes frontal e
parietal)
Beta 13 a 30 Hz Vigília. Estado de alerta.
Frontal (às
vezes occipital)
Gama 30 a 60 Hz
Intensa atividade mental; alta
concentra
ção; experiências
Diferentes áreas
corticais

82
místicas.
Lambda
Acima de
60 Hz
Estados extraordinários
(raros) de consciência; o
estado mais alto da meditação;
consciência plena de si
mesmo; introjeção profunda
(insight); ocasionalmente
quando se assiste TV;
estímulos visuais complexos.
Occipital
Ondas
Fusiformes
12 a 14 Hz
Está numa intersecção entre a
faixa alfa e a beta: estágio
inicial do sono.
Frontal e
parietal
Geralmente, consideram-se apenas quatro delas, por ordem de freqüência: delta,
teta, alfa e beta, que ocupam a faixa de freqüência entre 0.5 e 30 Hz.
“Quando o tálamo transmite ao córtex as informações provenientes dos
sistemas sensoriais, ou mesmo as que vêm de outras regiões, o número e a
variedade de potenciais sinápticos gerados são tão grandes que os eletródios
posicionados do lado de fora do crânio só conseguem captar a sua soma
algébrica que as aproxima de zero. O resultado é um traçado dessincronizado,
isto é, composto por ondas de baixa voltagem e alta freqüência (ritmos α e
β). É o que ocorre quando o individuo está acordado. Mas quando o tálamo
não deixa passar tão facilmente a informação que recebe, tornam-se menores,
menos variados e mais sincronizados os potenciais sinápticos no córtex.
Resulta um traçado sincronizado, composto por ondas de alta voltagem e
baixa freqüência (ritmos θ e δ)”. (LENT, 2001)
Um registro de EEG conterá todas as freqüências descritas na Tabela 2.1.
Entretanto, um tipo de onda terá maior ou menor predominância em função da região do
escalpo que está sendo analisada, bem como do estado do indivíduo, como vigília, sono,
coma, relaxamento; ou ainda, estados patológicos como epilepsia, comportamentos
psicóticos, demência, etc.
83
Figura 2.40 – Exemplos de ondas cerebrais (adaptada de (BUTTON, 2000)
& BERNARDI, 1999))
2.8 Epilepsia
A epilepsia é caracterizada pela atividade excessiva e descontrolada de uma
parte do sistema nervoso central, associada a crises focais, ou envolvendo todo ele,
associada a crises generalizadas.
O cérebro, em seu perfeito estado de saúde, apresenta ritmos elétricos de uma
forma ordenada. Assim, na epilepsia, essa ordem é alterada e surgem grupos de
neurônios com descarga desordenada, desobedecendo aos padrões de ondas cerebrais,
provocando verdadeiras “tempestades elétricas”.
As causas da epilepsia são bastante variadas: doenças parasitárias,
principalmente a neurocisticercose, outras infecções intracranianas de origem bacteriana
ou viral, traumas cranianos, tumores doença vascular, fatores hereditários e danos
cerebrais permanentes, como neurocisticercose, malária, meningite, desnutrição e
complicações perinatais, correspondem a uma das principais explicações para a maior
incidência de epilepsia nos países em desenvolvimento (PEREIRA, 2004).
A epilepsia pode ser classificada em três tipos principais segundo (GUYTON,
2002): a epilepsia do grande mal, a epilepsia do pequeno mal e a epilepsia focal.
A epilepsia do tipo grande mal é caracterizada por extrema descarga neuronal
em todas as áreas do cérebro: no córtex cerebral, nas partes mais profundas dos
hemisférios cerebrais, no tronco cerebral e no tálamo. Também, as descargas
transmitidas até a medula espinhal causam crises tônicas generalizadas no corpo inteiro,
seguidas mais ao fim do ataque, por contrações musculares alternadas tônicas e, depois,
espasmódicas denominadas de “crises tônico-clônicas”. Nesse tipo de crise a pessoa
pode cair no chão, apresentar contrações musculares em todo o corpo, ter perda súbita
da consciência, mordedura da língua, salivação intensa e respiração ofegante.
A epilepsia do tipo pequeno mal (“petit mal”) envolve o sistema ativador
cerebral talamocortical básico. É caracterizado geralmente por 3 a 30 s de inconsciência
ou diminuição da consciência, durante os quais a pessoa apresenta várias contrações
84
musculares em forma de abalos, geralmente na região da cabeça, sobretudo piscar dos
olhos; isto é seguido pela volta da consciência e a retomada das atividades prévias.
Muitas vezes não é percebida pelas pessoas, por ser curtíssima.
A epilepsia do tipo focal pode envolver praticamente qualquer parte do cérebro,
como as regiões localizadas do córtex ou estruturas mais profundas, tanto nos
hemisférios cerebrais quanto do tronco cerebral. Quase sempre, a epilepsia focal resulta
de alguma lesão orgânica localizada ou anormalidade funcional, como tecido neuronal
adjacente, um tumor que comprime uma área do cérebro, uma área de tecido cerebral
destruído, ou circuitos locais congenitamente perturbados.
Existem outros tipos de crises que podem provocar quedas ao solo sem nenhum
movimento ou contrações ou, então, induzir percepções visuais ou auditivas estranhas
ou, ainda, alterações transitórias da memória.
Esta patologia afeta 1% da população mundial, ou seja, 50 milhões de pessoas,
dos quais 25% não respondem a medicamentos anticonvulsivantes, que constituem a
terapia mais comum. Dentro os que recebem medicação, estima-se que 50% apresentem
efeitos colaterais. A cada ano, dois milhões de novos casos são relatados na literatura
médica (URL 10).
Não se conhece totalmente o mecanismo desencadeador das crises epilépticas. A
quantidade de pacientes epilépticos, cujo tratamento é atualmente sem sucesso ou que
sofrem de efeitos colaterais nocivos oriundos da medicação, exige a busca de soluções
alternativas, como por exemplo, novas técnicas de investigação informatizadas,
neurocirurgias e o desenvolvimento de medicamentos mais eficientes, e
consequentemente, caros. Assim sendo, nos países subdesenvolvidos, a terapia
medicamentosa pode chegar a ter um custo mais elevado do que nos EUA ou na Europa.
Considerando o baixo poder aquisitivo desses pacientes, e que o sistema público de
saúde (SUS) e os convênios não cobrem em geral estes tratamentos, mesmo as drogas
mais simples podem representar despesas proibitivas aos pacientes (COCKERELL,
1998).
IASEMIDIS (2003) discute a preditabilidade da crise epiléptica, supondo o
processamento de sinais EEG oriundos de diversos eletrodos, provendo informações
não obtidas pelo exame visual tradicional do EEG. O autor apresenta uma excelente
85
revisão bibliográfica acerca das diversas aproximações teóricas de análise do sinal EEG,
que podem ser resumidas na teoria de sistemas dinâmicos não-lineares, caos, análise
estatística através da filtragem adaptativa e de redes neurais. Estas pesquisas sugerem
estratégias para a construção de neuroimplantes, que minimizam as crises epilépticas,
conforme experimentos preliminares de eletro-estimulação realizados em animais
(LITT, 2003).
Ao mesmo tempo (NICOLELIS, 2002), as interfaces cérebro-máquina (ICM)
podem auxiliar na prevenção de crises cerebrais associadas à epilepsia crônica, que
podem ocorrer várias vezes por dia, diminuindo bastante a qualidade de vida do
paciente, além de possibilitar danos cerebrais permanentes. Estas interfaces
funcionariam como marcapassos cardíacos, monitorando a atividade elétrica cerebral e
identificando fatores desencadeantes de uma crise re-entrante. Uma vez detectada a
possibilidade iminente de crise, com razoável confiabilidade, o dispositivo provocaria
uma eletroestimulação no cérebro ou em um nervo periférico, ou ainda provocaria a
liberação de medicação antiepiléptica, com o objetivo de eliminar a crise.
Portanto, a epilepsia consiste em um importante problema de saúde pública, em
particular no Brasil, o que motiva estudos aprofundados e iniciativas governamentais
(MIN, 2003), no sentido de desenvolver novas técnicas para prevenção e tratamento da
doença.
2.9 Conclusão
Dentro do que foi exposto neste capítulo, realizou-se uma síntese dos diversos
conceitos em neurofisiologia. Enfocou-se o estudo no neurônio, visto que é o tema
principal nos tratados de neurociências, considerado o responsável pelo processamento
de sinais e pela memória. Ele também possibilita a transmissão do impulso nervoso
através das sinapses.
Dada à necessidade de medir a atividade elétrica celular, a ciência evoluiu do
“patch-clamp” para a criação de novas técnicas de medida neurofisiológica, através das
interfaces bioeletrônicas e, posteriormente, para a aquisição sistemática de sinais em
86
cultura através das matrizes multieletrodo. Esta revolução, restrita apenas a alguns
centros de pesquisa, é limitada pelos elevados custos da tecnologia MEA.
A epilepsia consiste num desafio da saúde pública mundial, em termos de
controle e prevenção. Atualmente, 60 milhões de pessoas são portadores da doença,
cujo tratamento é baseado em drogas anticonvulsivantes ou em cirurgias. Deste total,
aproximadamente 18 milhões de pacientes não conseguem obter um controle efetivo de
suas crises, sofrendo os danosos efeitos colaterais das terapias em uso.
A epilepsia pode ser considerada como fruto do desregulamento do conjunto de
atividades elétricas da massa neural que compõe o cérebro humano. Desta forma, a
interação simultânea de milhares de células nervosas, correspondente à mistura de
milhares de potenciais de ação, origina um padrão neurodinâmico patológico. Este
padrão elétrico é comumente mensurado através do EEG, que permite uma avaliação
extremamente grosseira do estado de ativação dos neurônios piramidais do córtex. Parte
do diagnóstico clínico da epilepsia, bem como de outros estados cerebrais patológicos,
se baseia nesta medida grosseira.
Por outro lado, as MEAs constituem a base tecnológica dos novos sistemas de
eletroestimulação e/ou neuroimplantes. Desta forma, a terapia através de
neuroimplantes (por exemplo, de retina e de cóclea) pode contar com novas
perspectivas, oriundas da nanotecnologia e de neuroimplantes auto-ajustáveis,
adaptados para cada paciente. Em particular, (LITT, 2003) descreveu planos para a
construção de neuroimplantes que minimizam crises epilépticas, de acordo com
experimentos em animais.
Desta forma, as MEAs tendem, em médio e longo prazos, a processar também
sinais EEG. Deve-se destacar que a restrição de biocompatibilidade exige o mínimo
consumo de potência por parte dos circuitos de neuroimplantes.
Em resumo, conclui-se que a utilização de novas terapias baseadas em
neuroimplantes na prática clínica estabelece os seguintes desafios para o processamento
de sinais:
(D1) Necessidade de se desenvolver novos enfoques teóricos e ferramentas,
adequados ao contexto não-estacionário dos sinais e à não-linearidade dos sistemas.
87
(D2) Necessidade do desenvolvimento de algoritmos simples e de baixa
exigência computacional (para minimizar a potência consumida e viabilizar a
biocompatibilidade), que permitam operação em tempo real.
(D3) A capacidade de processar simultaneamente, através de MEAs, tanto sinais
biológicos em nível celular (potenciais de ação, com objetivos fisiológicos), como em
nível neurológico (EEG, com objetivos clínicos). Em particular, o estabelecimento de
conexões entre as informações ligadas aos dois níveis de sinais permitirá uma melhor
compreensão do funcionamento do sistema nervoso humano.
Tendo em mente os desafios (D1) – (D3), encerra-se este capítulo com o
objetivo de se procurar, na literatura, técnicas de processamento de sinais para se
alcançar a viabilização de neuroimplantes como terapia.
88
Capítulo 3
Síntese Bibliográfica sobre Processamento de
Sinais EEG e MEA
3.1 Introdução
Este capítulo resume as principais tendências ligadas à abordagem da não-
estacionariedade em sinais biológicos, com especial enfoque aos sinais MEA. Uma vez
discutido alguns aspectos gerais do janelamento, detalham-se as principais técnicas de
processamento de dados oriundos de medidas intracelulares em culturas. A síntese
apresentada a seguir deve ser lida e ponderada à luz dos desafios (D1)-(D3)
estabelecidos no Capítulo 2.
3.2 Considerações gerais e processamento de sinais EEG
Aspectos importantes ligados a um sinal biológico qualquer correspondem à
não-linearidade do sistema envolvido ((MARMARELIS, 1978), (BARDAKJIAN,
2003)); bem como o fato das séries temporais serem não-estacionárias ((AKAY, 2003),
(MARMARELIS, 1994)). A não-linearidade está associada à complexidade,
característica dos organismos ((IASEMIDIS, 2003), (RIKE, 1997)); enquanto que séries
temporais não-estacionárias decorre de condições e circunstâncias peculiares de um ser
vivo, o qual deve sobreviver no meio em que está inserido (KANDEL, 2000).
89
A principal conseqüência prática de ser não-estacionária reside na dificuldade de
se dividir o arquivo de dados sob análise em pequenas partes (operação de janelamento
ou segmentação), considerando que, a princípio, não se têm conhecimento prévio da
variação temporal. Visto que qualquer tipo de processamento exige um janelamento
ótimo, o desenvolvimento de equipamentos e de métodos de análise quantitativa pela
engenharia biomédica fica condicionado ao desempenho da segmentação.
Pode-se considerar, na literatura, duas aproximações principais para se realizar o
janelamento em ambiente não-estacionário. A primeira ((COHEN, 1986), (GLASER,
1976)), considera apenas o domínio do tempo, dividindo o sinal biológico em trechos
estacionários. Desta forma, pode-se aplicar técnicas baseadas na hipótese de
estacionariedade. Por exemplo, a detecção de rupturas em modelos paramétricos
(COHEN, 1986), a segmentação adaptativa (HILTUNEN, 2000), e os modelos
autorregressivos variantes no tempo (GLASER, 1976). Estes métodos são
caracterizados (THAKOR, 2004) pela baixa complexidade computacional e
implementação adaptativa, o que permite operação em tempo real. Por outro lado, os
resultados podem ser incoerentes e imprecisos, devido à divergência dos métodos.
A análise de eletroencefalograma (EEG) se baseia, de acordo com a literatura,
em técnicas da primeira aproximação ((LITT, 2002), (MORMANN, 2003),
(MORMANN, 2004), (THAKOR, 2004)). Já para a codificação neural ((RIKE, 1997),
(BORST, 1999)), aplicam-se também ferramentas tradicionais, sem todavia se
aprofundar o aspecto de janelamento. Por exemplo, em (EICHLER, 2003), os autores
utilizam à função de correlação cruzada para estudar a conectividade neural, ou seja, a
rede de conexões morfológicas estabelecidas entre as células que compõem a cultura
celular sob análise. Com este mesmo objetivo, as densidades de probabilidade espaço-
temporais de sinais MEA são estimadas em ((VAN PELT, 2004), (CHIAPPLONE,
2003), (NOVELLINO, 2003)), enquanto (COZZI, 2005) utiliza uma aproximação do
tipo função de transferência.
A segunda aproximação para o estudo do janelamento em ambiente não-
estacionário leva em conta o domínio do tempo-freqüência, fundamentada na teoria de
wavelets ((STRATIMIROVIC, 2001), (SAMAR, 1999), (AKAY, 2003)); na análise de
flutuação aleatória ((BLESIC, 1999), (BLESIC, 2003)) e na teoria de sistemas

90
dinâmicos não-lineares ((BLANCO, 1995), (LITT, 2002), (MORMANN, 2003),
(MORMANN, 2004)). Estas técnicas apresentam alta complexidade computacional, e,
portanto, máxima precisão dos resultados, com convergência assegurada. Por outro
lado, a operação em tempo real fica condicionada à implementação de estruturas de
processamento especializadas e, consequentemente, mais complexas e caras.
3.3 Processamento de sinais MEA
3.3.1 Pré-processamento e geração da série temporal de intervalos
entre spikes
O primeiro estudo sobre a comunicação neural baseada nos sinais elétricos foi
realizado por Luigi Galvani (1791), quando demonstrou que os músculos da rã podiam
ser estimulados pela eletricidade. A Figura 3.1 ilustra a forma de onda de um sinal
extracelular, registrado eletronicamente. Nesta figura, o eixo horizontal corresponde ao
tempo analógico; enquanto que o eixo vertical representa a amplitude da tensão elétrica
absoluta, medida diretamente nos eletrodos do dispositivo de mensuração. A série
temporal representada pela Figura 3.1 será denominada “Atividade Neural Espontânea
Absoluta” (ANEA).
Figura 3.1 – Forma de onda extracelular com diferentes potenciais de ação, definindo a
série temporal ANEA (adaptada de (LEWICKI, 1998))
91
O sinal da Figura 3.1 apresenta vários picos ou spikes, por exemplo, nos
instantes de tempo 4.5, 5.1, 9.0 e 23.0; os quais quase sempre correspondem à
ocorrência de potenciais de ação. O processo de checagem desta correspondência é
denominado “detecção de spikes” (LEWICKI, 1998). Deve-se notar que, durante os
intervalos de tempo compreendidos entre a ocorrência de dois spikes consecutivos, o
sinal da Figura 3.1 apresenta amplitudes baixas, que oscilam em torno de um valor
médio. Isto pode ser observado nos intervalos 3-4, 6-8.9, 13-22. Estes trechos de sinal
são denominados “atividade basal” (KANDEL, 2000).
Do ponto de vista clássico ((KANDEL, 2000), (LEWICKI, 1998), (RIKE,
1997)), considera-se que a informação estatística fundamental de uma série ANEA está
concentrada nos spikes, ou melhor, nos instantes de tempo em que os spikes acontecem.
Consequentemente, a informação biológica ligada à atividade basal é descartada, sendo
que, na prática atual (RUTTEN, 2002), não se utiliza a série ANEA. De fato, com base
nesta última, os instantes de tempo ti em que os spikes acontecem são determinados,
gerando uma série do tipo {t1, t2, t3, t4, t5, t6, ...}. Em seguida, estimam-se os
intervalos de tempo entre spikes consecutivos, o que permite definir o sinal denominado
“série temporal de Intervalo Entre Spikes” (IES), definida logo abaixo.
{t2 - t1, t3 – t2, t4 – t3, t5 – t4, t6 – t5, ...}
A detecção do spike é uma técnica que permite classificá-los, de forma a
permitir o estudo da população local dos neurônios que estão isolados por técnicas
tradicionais (LEWICKI, 1998). Se a atividade dos neurônios pode ser uma característica
de um eletrodo, isto possibilita a classificação dos spikes, mesmo no caso de dois ou
mais neurônios conectados simultaneamente. Existem muitos problemas básicos ligados
ao processamento de spikes. Um deles é a detecção do ponto inicial, o qual depende da
amplitude da tensão elétrica absoluta, que pode ser observada no osciloscópio, mas com
a desvantagem de não se poder analisar o spike isoladamente.
Um outro método para análise de spikes em um longo período de tempo é o
histograma “intervalo entre spikes”, correspondente a uma estimativa da função
densidade de probabilidade da variável aleatória intervalo entre spikes. Embora simples,
este método requer um grande número de pontos confiáveis em um longo período de
tempo, tendo em vista a necessidade de uma quantidade mínima de dados que permita

92
estimadores estatísticos confiáveis. Consequentemente, o cálculo deste histograma não
ocorre em tempo real.
A eficiência de análise do spike é limitada pela detecção da altura do potencial
de ação. Para muitas situações, dois neurônios diferentes geram potenciais de ação com
formas de onda similares, durante o registro. Isto acontece quando os neurônios são
muito similares na sua morfologia e igualmente distantes do eletrodo que está sendo
usado para registro.
Em (CHAN, 2004), introduz-se o método baseado em população de spikes (PS),
representado pelas respostas dos neurônios monitorados com atividades extracelulares.
Essa resposta pode ser modelada pelo modelo Pulse-In-Pulse-Out. O objetivo é detectar
a ocorrência de spikes, cujas amplitudes, extraídas em tempo real, são necessárias para
implantes neurais ou próteses, porque modelos neurais computacionais usualmente
requerem uma entrada, expressa pela amplitude do impulso com determinada resposta.
Isto é, a extração da amplitude do spike consiste no primeiro passo para a análise
computacional dos neurônios. O artigo também apresenta uma implementação do
hardware para aproximação de cálculo de PS, que é um componente para pesquisas e
desenvolvimento de próteses dedicadas ao hipocampo cortical. A amplitude PS é
definida pela distância entre o spike mínimo e o ponto que corresponde à projeção do
mínimo na tangente linear entre dois picos. O algoritmo envolve a primeira diferença da
forma de onda. Essa operação de diferenciação é feita pelo cálculo da integral do sinal
no tempo real, que resulta nas amplitudes associadas. É utilizado um filtro de resposta
impulsiva infinita (IIR).
A fórmula matemática equivalente é definida por:
21121
0
)()()0()(././
2
1
1
pp
T
T
T
AATyTyyTydtdtdydtdtdy +=−+−=+
∫∫
(3.1)
Onde y representa o sinal de atividade neural espontânea no domínio do tempo, e os
termos da equação correspondem às amplitudes
e de acordo com a Figura 3.2.
1p
A
2p
A

93
Figura 3.2 - Métodos de extração da população dos spikes (PS)
(adaptada de (CHAN, 2004))
Logo, a altura desejada do PS pode ser aproximadamente calculada por:
2/)(
21 ppc
AAA
+
=
(3.2)
Ou seja,
representa a área média da função derivada primeira do sinal y(n),
no intervalo de tempo compreendido entre 0 e T
c
A
2
. Se esta área média for maior que um
determinado limiar, significa a ocorrência de uma variação brusca na amplitude de y(n),
característico dos mecanismos de polarização-repolarização ligados ao potencial de
ação. Decide-se então pela ocorrência do spike. Do contrário, durante o trecho
analisado, o sinal da MEA é considerado como atividade basal, que é caracterizada por
uma constância temporal na amplitude de y(n), o que se reflete como derivada
aproximadamente nula.
3.3.2 Classificação de spikes
Uma vez detectados os spikes, as séries temporais de intervalo entre spikes são
estimadas. Assim sendo, deve-se agrupar os diversos spikes em grupos semelhantes, o
que é realizado pelo procedimento de “classificação de spikes”.
O autor (STAVEREN, 2002) realizou testes para a caracterização da atividade
de potenciação (“spikes”) espontânea, modificada por testes de estimulação externa e de
treinamento. Os dados foram obtidos de neurônios corticais do cérebro de ratos

94
neonatais, durante um período de 4 horas. É utilizado o cloreto de sódio (NaCl) na
solução para não remover a aderência das células. A cultura média é realizada três vezes
por semana. O software quantifica e classifica as formas de ondas dessas atividades
espontâneas, onde a entrada do sinal é processada em tempo real, sendo que a redução
do ruído é feita via software. A atividade medida é dividida em canais, onde cada canal
assume um nível detector para excluir a atividade cíclica do sinal.
O autor utilizou o método de classificação da “distância mínima de
Mahalanobis”, onde primeiramente ele seleciona os spikes manualmente, em uma
pequena quantidade de classes. Durante a fase do treinamento, é representada a forma
de onda e suas características, através de uma seleção aleatória de spikes. Logo, os
spikes são classificados por classes, de acordo com a menor distância de Mahalanobis,
que é estimada a partir do cálculo da média e da covariância, de acordo com a seguinte
equação:
)(**)(
1
, i
t
iisMahalanobi
xxD
µµ
−−=
−
(3.3)
Onde
x é o vetor de spikes de amplitudes de tensão com valores entre 50 e 350 µV,
i
µ
é o vetor médio representativo da classe, ou seja, calculado pela mínima distância de
Mahalanobis; e
C é a matriz de covariância da classe.
Os resultados foram obtidos em duas experiências com culturas de ratos
embrionários, considerando aproximadamente 180.000 spikes. A classificação é feita
em clusters que representam seis formas de onda básicas, com suas respectivas
morfologias.
Em outro artigo, o autor (IZHIKEVICH, 2003) apresenta o modelo dos
neurônios do tipo “spiking” (potenciação) e “bursting” (salva), baseando-se na dinâmica
do tipo de Hodgkin-Huxley de duas dimensões, canônico, utilizando equações
diferenciais ordinárias da seguinte forma:
Iuvv
dt
dv
+−++= 140504,0
2
(3.4)
)( ubva
dt
du
−=
(3.5)

95
Com algumas restrições: se
entãomVv ,30≥
duu
cv
+→
←
Onde v representa o potencial da membrana do neurônio, o termo 0,04v
2
+5v+140
representa o ajuste dinâmico dos picos e a variável u representa o potencial de
recuperação da membrana, relativamente aos íons Na
+
e K
+
, gerando valores negativos
para v.
As variáveis a, b, c e d são parâmetros, onde a corresponde ao tempo adquirido
da variável u, e assume o valor de a = 0,02. Já o parâmetro b é a descrição da variável u
em relação ao potencial da membrana v, assumindo o valor b = 0,2. O parâmetro c é o
valor da condutância do íon K
+
, no potencial de membrana, e possui o valor de c = -65
mV. O parâmetro d corresponde à amplitude das condutâncias dos íons Na
+
e K
+
, com
valor d = 2. Durante a implementação, são simulados em tempo real milhares de
neurônios conectados, explicitando as áreas inibitórias e as excitatórias da rede neural.
Todas as células corticais excitatórias são divididas em quatro classes e todas as células
corticais inibitórias são dividas em duas classes, de acordo com os tipos padrões de
spikes.
Muitas técnicas de classificação de spikes para eletrodos simples se estenderem
naturalmente aos eletrodos múltiplos (LEWICKI, 1998). Além da análise de
componentes principais, pode-se também cogitar a aplicação da teoria de separação de
fontes (AKAY, 2003) para a situação de múltiplos canais, obtendo-se assim as
características de agrupamento. Neste caso, os componentes principais ou as funções
discriminantes descrevem a variação máxima direcionada para todos os canais
simultaneamente.
Finalmente, com base nas discussões apresentadas, pode-se concluir que a
classificação de spikes exige técnicas de reconhecimento de padrões, as quais, por sua
vez, não levam necessariamente em conta a operação em tempo real.
3.3.3 Estimação da conectividade neural
Em (CASTELLONE, 2003), o autor utiliza o histograma de intervalo de
neurônio cruzado (vide Figura 3.3), que analisa a conectividade entre pares de neurônios
da atividade espontânea em rede de cultura de células corticais de ratos.

96
Figura 3.3 - Histograma de intervalo de neurônio cruzado. Eixo y: intervalo
entre spikes [ms]. Eixo x: tempo [ms] (adaptada de (CASTELLONE, 2003))
Os histogramas mostram o intervalo dos spikes médio de acordo com suas
correlações entre os neurônios em vários períodos de tempo. O eixo x está dividido em
intervalos de tempo de 0,5 ms; enquanto o eixo y representa os tempos de intervalos de
spikes. A atividade espontânea é analisada pelo spike no tempo e pela localização dos
canais em atividade durante um período de 10 minutos. Quando o valor de AR
(amplitude) é baixo, indica a não-conectividade entre os eletrodos analisados.
Foram analisados 38 histogramas com 2 parâmetros: um de acordo com a forma,
e o outro de acordo com o intervalo de pontos. O histograma indica a (não) abundância
da conectividade e a conexão direta dos caminhos. Um caso especial, para a mesma
cultura, ocorre nas gravações entre 7 e 14 segundos, onde se observa um grande pico
entre os eletrodos. Os resultados foram similares àqueles obtidos pela literatura
((AERTSEN, 1985), (MELLSEN, 1987)), a partir da análise da correlação cruzada, para
gravação in vitro de cérebros animais.
Em (BORST, 1999), a codificação neural é considerada como a representação
neural da informação, em termos da teoria de probabilidade aplicada à transmissão de
informações em sistemas de comunicação. A teoria requer que as transferências das
informações sejam precisas, determinadas por distribuições de probabilidade exatas
particulares, para sinais de entrada e de saída. Neste contexto, o estudo da conectividade
neural corresponde a topologia que maximiza o fluxo de informação.
97
Por fim, nas referências ((EICHLER, 2003), (VAN PELT, 2004)), os autores
estudam a conectividade neural a partir da estimação da função de correlação cruzada
entre os sinais dos diversos eletrodos, sem contudo discutir explicitamente a operação
em tempo real do método, e nem o janelamento do conjunto de sinais levando em conta
a não-estacionariedade.
3.4 Conclusão
Dentre as duas principais aproximações para o janelamento de sinais não-
estacionários, a primeira delas, no domínio do tempo, parece melhor atender às
exigências de baixa complexidade computacional, visando a mínima dissipação da
potência e operação em tempo real, estabelecidas pelo contexto aplicativo de
neuroimplantes.
A pesquisa sobre codificação neural envolve diversas aproximações diferentes,
tais como processos de ponto (COHEN, 1986), modelos de caminhada randômicos
((GERSTEIN, 1964), (RIKE, 1997)) e análise do ruído branco (BLESIC, 1999). Todas
essas aproximações se baseiam uma série temporal obtida após a detecção e
classificação dos spikes, denominada “série de intervalo entre spikes”. Este sinal
apresenta diversas limitações e inconvenientes no contexto dos neuroimplantes, a saber:
A necessidade de detecção e classificação dos picos do sinal de atividade neural
espontânea. Observe que tal pré-processamento é inevitável, e ele é sujeito a
diversos erros devido o ruído que pertuba frequentemente a instrumentação
neurofisiológica (RUTTEN, 2002). Além disso, sempre existem incertezas a
respeito do limiar de amplitude a ser considerado, para se determinar se um pico
ocorreu em algum lugar ou não (RIKE, 1997). Consequentemente, tudo isso
dificulta a implementação do processamento de sinais em tempo real, através do
aumento da complexidade dos algoritmos; além de exigir o uso de técnicas de
filtragem de ruído, que necessariamente aumentam a potência dissipada pelo
circuito físico de aquisição dos sinais neurais.
A informação biológica ligada à atividade basal é perdida, o que pode justificar,
de certa forma, os limites de desempenho dos métodos clássicos.
98
Deve-se lembrar que, pela análise da literatura, os conceitos de histograma de
intervalo entre spikes, função de correlação cruzada; bem como as teorias de informação
e reconhecimento de padrões fundamentam a classificação dos spikes e o estudo da
conectividade neural. A leitura cuidadosa dos artigos consultados permite afirmar que
pouco se discutiu a respeito da operação em tempo real dos métodos, bem como do
janelamento dependente da não-estacionariedade dos sinais analisados.
Retornado agora aos desafios do Capítulo 2, fica claro a necessidade de, se
possível, utilizar diretamente a série temporal de atividade espontânea, ao invés da série
de intervalo entre spikes. Neste contexto, também seria desejável reduzir a
complexidade do hardware, substituindo um filtro de ruído aditivo por uma solução em
software.
Do nosso conhecimento, poucos trabalhos na literatura discutem as idéias
levantadas no parágrafo anterior, em particular, sobre a aplicação da teoria de
estatísticas de ordem elevada. Em (BOHTE, 2000), os autores apresentam uma análise
baseada em correlações de 3ª ordem, aplicadas a séries temporais do intervalo entre os
spikes. A gaussianidade ou não das séries temporais envolvidas, do nosso
conhecimento, não foi verificada.
Baseado nas motivações precedentes, o artigo (DESTRO-FILHO, 2001)
analisou a ligação entre a plasticidade sináptica e a filtragem de predição não-linear. Em
((BLESIC, 1999), (DESTRO-FILHO, 2002), (DESTRO-FILHO, 2003)), discutiu-se
também as propriedades do ruído branco da série temporal de intervalos entre spikes.
Resultados preliminares motivam o aprofundamento desta pesquisa.
Deve-se ressaltar a existência de dois resultados experimentais relativamente ao
janelamento de sinais EEG e de sinais de atividade neural espontânea registrados
através das MEAs. Tanto para o primeiro caso ((MORMANN, 2004),
(D’ALESSANDRO, 2005)), como para o segundo ((RAUCH, 2003), (BLESIC, 1999),
(BLESIC, 2003)), os autores enfatizaram a necessidade de particionamento dos sinais
em trechos de, aproximadamente, 2 segundos. Estas conclusões são essencialmente
experimentais, e não foram justificadas nem do ponto de vista de ciências exatas, nem
do ponto de vista fisiológico.
99
As conclusões acima apresentadas, bem como as interrogações do parágrafo
anterior, sugerem então o uso de técnicas simples para o processamento de sinais MEA
e EEG em tempo real, se possível evitando a detecção dos spikes. Uma análise
preliminar sugere a utilização da predição linear adaptativa, metodologia simples e bem
estabelecida, especialmente concebida para o processamento de sinais não-estacionários
no domínio do tempo. Enfim, a teoria de estatística de ordem elevada permite cogitar a
redução do ruído aditivo, freqüentemente encontrado em sinais neurológicos e
neurofisiológicos, através de técnicas de software. As duas teorias sugeridas são
aprofundadas no próximo capítulo.
100
Capítulo 4
Predição Linear Adaptativa e Estatística de
Ordem Elevada (EOE)
4.1 Introdução
No Capítulo 2 constatou-se a necessidade de utilização de algoritmos simples
para permitir o processamento, em tempo real, de sinais MEA e EEG através de
neuroimplantes. No Capítulo 3, concluiu-se que se pode fazer diversas aproximações
teóricas para a análise dos sinais de EEG e MEA: teoria de sistemas dinâmicos não-
lineares (HAYKIN, 2000), caos (DURAND, 2001), wavelets (TEOLIS, 1998) e redes
neurais (IASEMIDIS, 2003).
Neste capítulo, apresentam-se ferramentas teóricas que tentam, de uma certa
maneira, minimizar as limitações das técnicas atualmente utilizadas na análise da
codificação neural e da epilepsia (Capítulo 3), buscando atender ao máximo os desafios
especificados na conclusão do Capítulo 2. Propõe-se então o uso da filtragem adaptativa
e das técnicas de EOE para, respectivamente, lidar com os aspectos não-estacionários
dos sinais MEA/EEG, e com o caráter não-linear dos sistemas associados a estes
mesmos sinais.
A filtragem adaptativa constitui atualmente uma ferramenta imprescindível ao
processamento estatístico de sinais, principalmente daqueles resultantes de sistemas
cuja caracterização estatística é desconhecida. Assim sendo, o uso destes filtros permite
101
novas capacidades de processamento. Atualmente, esta técnica é aplicada com sucesso
em campos tão diversos como as comunicações, o controle, o radar, a sismologia e a
engenharia biomédica (HAYKIN, 1991).
Os filtros adaptativos são projetados de forma automática, através de um
algoritmo recursivo, tornando possível a operação satisfatória em ambiente não-
estacionário, onde o conhecimento das características dos sinais a processar não é
completo. O algoritmo se inicia com um conjunto de valores pré-determinados. No
entanto, após um determinado número de iterações, espera-se que o filtro convirja para
a solução ótima de Wiener. Em ambientes não-estacionários, o algoritmo permite ao
filtro a possibilidade de acompanhamento das variações estatísticas, desde que estas
sejam suficientemente lentas.
As estatísticas de ordem elevada, embora conceitualmente definidas e utilizadas
pela teoria de processos estocásticos desde o final do século XIX (NIKIAS, 1993),
foram aplicadas ao processamento de sinais a partir dos anos 1980-1990. Elas permitem
migrar do contexto clássico de análise, baseada nos sinais gaussianos e sistemas
lineares; para um enfoque mais genérico, onde os sistemas podem ser não-lineares, e os
processos estocásticos, não-gaussianos.
O capítulo discute, primeiramente, a filtragem adaptativa, suas aplicações, o
filtro de Wiener, e consequentemente, a Predição Linear Direta e Reversa, baseada no
modelo auto-regressivo (AR). Em seguida, abordam-se temas em EOE, como a função
característica, os cumulantes e também os índices de descorrelação.
4.2 Filtragem Adaptativa
A idéia de filtragem adaptativa emerge de um contexto onde os parâmetros do
filtro são obtidos a partir de operações matemáticas sobre o sinal que está sendo
processado.

102
Presume-se que o sinal tenha características variantes no tempo e o filtro deverá
sofrer constantes correções em seus parâmetros, de forma a se adaptar às novas
condições do sinal.
O termo “filtro” é utilizado para descrever um dispositivo em hardware ou de
software que se aplica a um conjunto de dados contaminados com ruído, de forma a
extrair a informação de interesse. O filtro é denominado linear se a quantidade de
interesse à saída do sistema é uma função linear das observações apresentadas a sua
entrada.
Considere a Figura 4.1 representando o diagrama de blocos do problema em
questão. O filtro tem como entrada o sinal desejado, contaminado com ruído branco
aditivo descorrelacionado, constituindo a seqüência x(0), x(1), x(2), ... e caracteriza-se
pela resposta impulsional w0, w1, w2, ... . Num determinado instante i, o filtro produz a
saída o(i). Esta é usada como estimativa da resposta desejada d(i). Sendo os dois sinais
oriundos de processos estocásticos, a estimativa produz um sinal de erro e(n) com
características estatísticas próprias. O objetivo é minimizar o erro de estimativa segundo
algum critério estatístico.
Figura 4.1 - Diagrama de blocos representando o problema de filtragem estatística (WIDROW, 1985)
A filtragem adaptativa tem sido aplicada com bastante sucesso em campos
diversos como comunicações, radar, sismologia e engenharia biomédica. Sendo
aplicações de natureza tão diferentes, todas partilham uma característica comum: um
vetor de entrada e uma resposta desejada são usados para calcular um erro de
estimativa, que, por sua vez, é usado para controlar um conjunto de coeficientes de um
filtro. Algumas das aplicações da filtragem adaptativa encontram-se descritas na Tabela
4.1.

103
Tabela 4.1 - Algumas aplicações da filtragem adaptativa.
Classe Aplicações
I. Identificação Identificação de sistemas
Modelização das camadas terrestres
II. Modelamento Inverso Desconvolução preditiva
Equalização adaptativa
III. Predição Codificação preditiva linear
IV. Cancelamento de
Interferências
Cancelamento adaptativo de ruído
Cancelamento de eco
1. Identificação: Utiliza–se um filtro adaptativo para obter o modelo que melhor
representa, segundo algum critério, o sistema desconhecido. Com isso, o sistema que se
pretende identificar e o filtro adaptativo são excitados com o mesmo sinal de entrada,
como esquematizado na Figura 4.2. O sistema desconhecido disponibiliza a resposta
desejada para o filtro adaptativo. Refira-se ainda que se o sistema for de natureza
dinâmica o modelo será também variante no tempo.
Figura 4.2 - Filtro Adaptativo usado para a identificação de um sistema (adaptada de (HAYKIN, 1991))
2 .Modelamento Inverso: Nesta classe, o filtro adaptativo é usado para obter o modelo
inverso de um sistema desconhecido. Numa situação ideal o modelo inverso representa

104
a função de transferência inversa da que caracteriza o sistema desconhecido. A versão
atrasada do sistema desconhecido representa a resposta desejada para o filtro adaptativo.
Figura 4.3 - Aplicação da filtragem adaptativa ao modelamento inverso (adaptada de (HAYKIN, 1991))
3. Predição: A função do filtro adaptativo consiste em disponibilizar a melhor predição
(segundo algum critério estatístico) do valor presente de um sinal aleatório. O valor
presente do sinal serve de resposta desejada para o filtro adaptativo. Note-se que o sinal
de referência do filtro consiste apenas nos valores passados do processo. Em alguns
casos, o sinal de saída do filtro adaptativo serve como saída do sistema, dizendo-se que
funciona como preditor. Em outros casos, o sinal de erro de predição é usado como
saída, afirmando-se nesta situação que o filtro funciona como um filtro de erro de
predição (prediction-error filter).
Figura 4.4 - Filtragem adaptativa aplicada à predição. A saída 1 corresponde à predição do
valor de entrada. A saída 2 refere-se ao erro de predição ( adaptada de (HAYKIN, 1991))
4.Cancelamento de interferência: Aqui o filtro adaptativo é usado para cancelar uma
interferência desconhecida que contamina o sinal de informação pretendido. O sinal de

105
informação com contaminação (sinal primário) é considerado como resposta desejada
para o filtro adaptativo. O sinal auxiliar, denominado de referência, é usado como
entrada para o filtro adaptativo. O sinal de referência obtém-se a partir de um sensor ou
conjunto de sensores colocados por forma a que contém a informação desejada seja
recebido em más condições, relativamente à interferência.
Figura 4.5 - Filtro adaptativo utilizado no cancelamento de interferências
(adaptada de (HAYKIN, 1991))
4.3 Cálculo dos Filtros
Filtros são utilizados com o objetivo geral de separar elementos diferentes.
Quando tratamos de sinais elétricos, os filtros são dispositivos que alteram o conteúdo
de freqüências do sinal de entrada, afim de remover partes indesejadas, por exemplo,
ruídos ou separar um sinal de outro.
4.3.1 Filtro Ótimo: aproximações de Wiener
Na década de 1940, Norbert Wiener foi pioneiro na pesquisa para a elaboração
de um filtro que produziria a estimativa ótima de um sinal ruidoso. Exibimos o
problema do filtro de Wiener, com objetivo de recuperar um sinal desejado, d(n), de
uma observação x(n) com ruído v(n) (vide Figura 4.6).
x(n) = d(n) +v(n)
(4.1)

106
Figura 4. 6 – Ilustração do problema geral do Filtro de Wiener. Dados dois processos estacionários,
x(n) e d(n), que são estatisticamente descorrelacionados entre si, o filtro W(z) minimiza a estimativa do
erro médio quadrático,
(n), de d(n). (adaptada de (DE FATTA, 1988)) d
ˆ
Assumindo que d(n) e v(n) são processos aleatórios estacionários e
estaticamente independentes, a elaboração do filtro consiste em minimizar o valor
esperado do erro médio quadrático da estimativa de d(n). Assim, como:
£ =
{
}
)²(neE
(4.2)
sendo:
E(n) = d(n) –
(n)
^
d
(4.3)
o problema é encontrar o filtro que minimize £.
Considerando o vetor formado pelo valor atual e anteriores de entrada no
instante i, temos:
[
T
Nixixix )1(...)1()() (i +−−=x
]
(4.4)
Designando por
R a matriz quadrada de autocorrelação do vetor de
entrada,
(4.5a)
⎪
⎪
⎭
⎪
⎪
⎬
⎫
⎪
⎪
⎩
⎪
⎪
⎨
⎧
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
+−−+−+−
+−−−−
+−−
==
)1²(...)1()1()()1(
............
)1()1(...)1²()()1(
)1()(...)1()()²(
](i)(i)[
NixixNixixN
ix
Nixixixixix
Nixixixixix
EE
T
xxR

107
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
−−
−
−
=
)0(...)2()1(
............
)2(...)0()1(
)1(...)1()0(
rNrNr
Nrrr
Nrrr
R
(4.5b)
onde r(k) é a autocorrelação de índice k = 0,1,2,...,N-1 do sinal x(n).
Sendo
P o vetor de correlação cruzada entre o sinal observado e o desejado:
[]
T
NixidixidixidEixidE 1()(...)1()()()()]()([ +−−==P
(4.6)
e considerando ainda que o filtro possui uma estrutura transversal com N elementos de
atraso, como ilustrado na Figura 4.7 , a sua saída o(i) será obtida pela expressão:
)()(
1
0
Nnxwio
N
n
n
−=
∑
−
=
(4.7)
Tal como em (4.3), pode-se considerar o vetor formado pelos coeficientes do
filtro:
[
]
T
N
www
110
...
−
=W
(4.8)
Figura 4.7 - Filtro Transversal com N coeficientes (adaptada de (HAYKIN, 1991))
obtém-se então a saída do filtro em notação matricial:
Wx
T
(i))( =io
(4.9)
Com isso pretende-se reduzir os efeitos do ruído à saída do filtro de acordo com
algum critério estatístico. Uma aproximação para este problema é minimizar o erro

108
quadrático médio do sinal do erro. Definindo sinal de erro como a diferença entre a
resposta desejada d(i) e o resultado obtido o(i),
)()()( ioidie
−
=
(4.10)
Com isso, o erro quadrático médio será:
[]
[
]
WPRWW
T
2)²()²( −+=
T
idEieE
(4.11)
A superfície do erro é um parabolóide multidimensional com concavidade
orientada no sentido positivo, ou seja, uma superfície com um único mínimo. Para
determiná-lo, calcula-se o gradiente da função:
PRW 22
)]([
2
−=
∂
∂
=∇
W
ieE
(4.12)
O vetor de pesos ótimo
W* é aquele para o qual se anula o gradiente:
0
)]([
2
=
∂
∂
=∇
W
ieE
(4.13)
Para sinais de entrada estacionários a solução resulta no vetor de coeficientes
W*, solução conhecida como filtro de Wiener e que será ótima no sentido quadrático
médio:
W* = P
R
1−
(4.14)
O desenvolvimento da (4.14) supõe as seguintes hipóteses:
(H1) Sinal x(n) estritamente estacionário, sem componente senoidal pura.
(H2) Ruído v(n) desprezível atuando sobre x(n), ou relação sinal-ruído
(SNR) elevada.
(H3) Filtro linear, adaptativo, inerentemente estável.
109
4.3.2 Filtros subótimos: aproximação estocástica
O filtro de Wiener, é, no entanto, inadequado para situações em que não existe
estacionariedade, requerendo conhecimento à priori da estatística dos dados a processar.
Este será ótimo se as estatísticas dos dados de entrada coincidirem com as que se
usaram no desenho do filtro. Quando não se dispõe dessa informação poderá não ser
possível desenhar o filtro de Wiener, ou, pelo menos, não terá um desempenho ótimo.
Uma aproximação expedita nesta situação poderá consistir na separação do
processamento em duas etapas: na primeira estimam-se os parâmetros estatísticos dos
sinais e na segunda, aplica-se ao cálculo dos parâmetros do filtro. Nas aplicações em
tempo real, este processo poderá ser muito dispendioso, devido às exigências
computacionais.
Sendo múltiplos algoritmos existentes, dividem-se, no entanto em apenas duas
classes. A primeira são os Algoritmos de Gradiente e a segunda os algoritmos de
mínimos quadrados.
Os algoritmos de gradiente baseiam-se no filtro de Wiener, estimando o
gradiente da superfície da função de custo. Tem como principal vantagem à baixa
complexidade. São bons na exploração de superfícies pouco complicadas.
Os algoritmos de mínimos quadrados são baseados no filtro de Kalman. De
forma determinística minimizam a soma dos quadrados dos erros parciais. Tem como
vantagem a baixa sensibilidade a mínimos locais das superfícies da função de custo e a
maior velocidade de convergência comparando-o com os algoritmos de gradiente.
Dentre as desvantagens, estão as maiores exigências computacionais e problemas de
estabilidade.
Em seguida, discute-se detalhadamente o algoritmo dos mínimos quadrados
médios (“Least Mean Square” - LMS), tendo em vista sua baixa complexidade
computacional, o que é uma característica fundamental para assegurar a
biocompatibilidade de um neuroimplante.
Este algoritmo é importante pela sua simplicidade e facilidade de computação.
Se o sistema adaptativo é um combinador linear adaptativo, e o vetor de entrada x(n) e a

110
resposta desejada d(n) estão disponíveis a cada iteração, o algoritmo LMS é geralmente
a melhor escolha para muitas aplicações de processamento de sinais (WIDROW, 1985).
Este algoritmo utiliza o valor instantâneo de e²(i) como estimativa da função de
custo calculada por E[e²(i)]. O algoritmo é o seguinte:
Passo1: Estima-se o gradiente da função:
[]
(i))(2
)(
)(i)(
2
^
x
W
eE
2
ie
ie
−=
∂
∂
=∇
(4.15)
Passo2: Como o gradiente é o vetor que aponta no sentido do máximo da função de
custo, desloca-se o vetor de pesos na direção oposta com o objetivo de procurar o
mínimo:
[
]
(
)
(i)e(i)1)(i
2
^^
EWW
^
∇−=+
µ
(4.16)
ou seja:
(i))(2(i)1)(i
xWW
^^
ie
µ
+=+
(4.17)
A constante µ designa-se habitualmente por passo de adaptação e controla a
estabilidade e velocidade de convergência do algoritmo. A existência de estabilidade do
algoritmo desde que o valor do passo de adaptação obedeça à seguinte desigualdade
(HAYKIN, 1991):
{}
))((*)1(
1
0
2
ixEN +
<<
µ
(4.18)
Logo, o erro residual será proporcional a µ, sendo por isso normal escolher
valores pequenos para esta constante.
Em situações de não-estacionariedade o passo muito reduzido levará a baixa
capacidade de seguimento das mudanças no ambiente de trabalho. Este algoritmo LMS
é atualmente o mais utilizado devido à sua baixa complexidade.
111
4.4 Predição Linear Direta (“forward”)
A modelagem paramétrica (HAYKIN, 2000) é uma técnica para análise de séries
temporais, na qual um modelo matemático é adaptado a um sinal amostrado. Se a forma
do modelo é uma boa aproximação para o comportamento observado do sinal, então
pode ser usado em uma ampla faixa de aplicações, como estimação espectral,
compressão de dados, síntese de fala, e extração de característica para classificação de
padrões.
Em modelos adaptativos, os valores dos parâmetros livres são atualizados com a
chegada de cada nova amostra de dados, considerando que em modelos não-adaptativos
os parâmetros são escolhidos de forma a melhor se ajustar a uma dada seqüência de
amostras.
A maioria dos sinais, inclusive a fala e o eletroencefalograma (EEG), são não-
estacionários (isto é, eles têm um espectro de freqüência variável no tempo), muito
embora eles possam ser considerados localmente estacionários para intervalos de tempo
pequenos.
Para tais sinais, qualquer modelo adaptativo pode ser usado, ou então o sinal
pode ser dividido em segmentos suficientemente pequenos, quase estacionários, e um
modelo não-adaptativo deve ser ajustado para cada segmento. Técnicas de modelagem
paramétricas são usadas para calcular os parâmetros livres nos vários algoritmos
existentes.
Os algoritmos rápidos exploram a natureza seqüencial do sinal de entrada. Ou
seja, o vetor
x(n) pode ser obtido, a menos do primeiro elemento, por um simples
deslocamento nos elementos de x(n-1). Se existe correlação entre as amostras, é também
possível fazer uma estimativa do primeiro elemento de x(n) a partir das amostras
anteriores.
Esta estimativa é tanto melhor quanto maior for a correlação existente entre as
amostras. O processo de estimar as amostras do sinal é denominado de predição.
112
4.4.1. Modelo Auto-Regressivo (AR)
Fazemos um modelamento do sinal a partir de parâmetros estabelecidos. A
entrada do modelo corresponde a um sinal de ruído branco, por exemplo, que pode ser
facilmente gerado de forma computacional através do gerador de números aleatórios do
MATLAB. A saída do modelo, idealmente, corresponderia a um sinal captado de um
eletrodo, que gera sinais biológicos.
Matematicamente, existem diversos modelos para se relacionar a variável x(n) a
outras variáveis nos quais queremos analisar. A técnica de predição linear considera
uma relação linear, conforme especificado abaixo.
x(n) = a
0
v(n) + a
1
v(n-1) + ... + a
n-1
v(n-N+1) (4.19)
x(n) =
)(
1
0
inva
N
i
i
−
∑
−
=
(4.20)
onde { a
0
, a
1,
..., a
N
–1
} são coeficientes (constantes) e N é a ordem do modelo.
4.4.2 Modelo da Predição Linear Direta
Neste modelo consiste em prever uma amostra de sinal a partir de suas amostras
analisadas anteriormente.
)()1(
1
0
^
jnxanx
N
j
j
−=+
∑
−
=
(4.21)
)1(...)1()()1(
110
^
+−++−+≅+
−
Nnxanxanxanx
N
(4.22)
onde o valor de x previsto (
) é calculado a partir dos xs já conhecidos.
^
x
Para definir se o modelo utilizado está adequado, analisamos a diferença entre o
sinal previsto e o sinal real. Essa diferença é denominada “erro de predição”.
)()()(
^
nxnxne
p
−=
(4.23)
113
onde e
p
é o erro de predição e (n) é calculado a partir de : { x(n-1), x(n-2),....,x(n-
N)}.
^
x
4.4.3 Critérios para uma boa predição
Determinamos alguns critérios para que esse modelo esteja funcionando de
forma adequada. Por exemplo:
1- e
p
(n) → 0
2-
→ 0
{
)(
2
^
neE
p
}
Com isso, o erro de predição dever tender a zero e a esperança do erro de
predição ao quadrado deve, também, tender a zero. O segundo critério é extremamente
importante, tendo em vista a natureza estocástica e não-estacionária dos sinais
biológicos envolvidos e analisados como os dados de entrada.
4.4.4 Solução ótima de Wiener para Predição Linear Direta
Para resolvermos as equações anteriores a partir das Equações de Wiener-Hopf
temos três características importantes. A primeira relacionada com matriz de correlação
de ordem N por N com os valores de entrada {x(n-1), x(n-2),...,x(n-N)}. A segunda,
relacionada ao vetor de correlação cruzada de ordem N por 1, entre as entradas e a saída
desejada x(n). O valor do erro de ordem N, é estimado como a terceira característica, a
variância de x(n).
1) Os valores de entrada {x(n-1), x(n-2), ...., x(n-N)} são armazenados no seguinte
vetor:
T
Nnxnxnx ](...)2()1([1)(n −−−=−x
(4.24)
Portanto, a matriz de correlação dos valores de entrada será:
]1)(n1)(n[ −−=
T
xxR E
114
=
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
−−
−
−
)0(....)2()1(
..............
)2(....)0()1(
)1(....)1()0(
rNrNr
Nrrr
Nrrr
(4.25)
Podemos observar que o símbolo usado na matriz de correlação para os valores
de entrada é semelhante aos valores da matriz de correlação do filtro transversal da
Figura 4.7.
2) O vetor de correlação cruzada entre os valores de entrada {x(n-1),....x(n-N)} e a
saída desejada x(n) é:
)](1)(n[ nxE
−
=
xrp
=
=
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
)(
.....
)2(
)1(
Nr
r
r
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
−
−
−
)(
...
)2(
)1(
Nr
r
r
(4.26)
3) A variância de x(n) é igual a r(0), desde que x(n) tenha média zero.
De acordo com (4.4)-(4.6), nós adaptamos as equações de Wiener para resolver
o problema de predição linear direta para entradas estacionárias, onde:
R a = rp
(4.27)
Similarmente, usando (4.14) e (4.26), deriva-se a seguinte expressão do erro de
predição:
arp
T
−= )0(re
p
(4.28)
Para as equações anteriores, tem-se um vetor de pesos Nx1, determinados
somente por um conjunto de (N+1) valores da função de autocorrelação.
Exemplo 4.1: Processo estocástico associado a um ruído de 60 Hz, presente em
sinais de matriz multieletrodo (MEA) (LATHI, 1967).
Seja um processo estocástico definido por:
)cos((t)
φ
ω
+
=
tAx
(4.29)

115
onde
φ
é uma variável aleatória com distribuição uniforme entre 0 e 2π.
Conforme demonstrado em LATHI (1967), temos que :
)cos(
2
)(
2
ωττ
A
R
x
=
(4.30)
A matriz de autocorrelação, de acordo com (4.5a)-(4.5b), considerando
τ
=0,1,
será:
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
=
2
)cos(
2
)cos(
22
22
22
AA
AA
ω
ω
R
(4.31)
Logo, o vetor de correlação cruzada para
τ
=1,2 é:
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
=
)2cos(
2
)cos(
2
2
2
ω
ω
A
A
p
(4.32)
De acordo com as equações de Wiener, descritas anteriormente, e usando (4.14),
calcula-se o vetor de coeficientes a partir de:
⎥
⎦
⎤
⎢
⎣
⎡
⎥
⎦
⎤
⎢
⎣
⎡
−
−
−
=
)2(
)1(
*
)0()1(
)1()0(
*
)1()0(
1
22
r
r
rr
rr
rr
w
(4.33)
Substituindo (4.30) na equação anterior, obtém-se:
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎣
⎡
−−
−
=
)2cos(
)(cos
)2cos()cos()cos(
44
2
44
ω
ω
ωωω
AA
AA
w
(4.34)
Logo, considerando A=2 e f=60,1 Hz, e utilizando o programa MATLAB para o
cálculo, teremos os seguintes resultados:
⎥
⎦
⎤
⎢
⎣
⎡
=
26180,1
6180,12
R
⎥
⎦
⎤
⎢
⎣
⎡
=
6180,0
6180,1
p
⎥
⎦
⎤
⎢
⎣
⎡
−
=
1
6180,1
w
Consequentemente, para A=2 e f=1000,2 Hz, teremos:

116
⎥
⎦
⎤
⎢
⎣
⎡
=
26180,0
6180,02
R
⎥
⎦
⎤
⎢
⎣
⎡
−
=
6180,1
6180,0
p
⎥
⎦
⎤
⎢
⎣
⎡
−
=
1
6180,0
w
Analisando estes resultados, percebemos que são semelhantes, mudando apenas
a freqüência do sinal. Deve-se destacar que w(2) é constante, assumindo o mesmo valor
nos dois casos.
As Figuras 4.8 e 4.9 ilustram os coeficientes do vetor w obtidos em vários testes,
de acordo com (4.34), considerando A=2 e a freqüência variando de 60 Hz a 1 kHz.
Para cada gráfico, mostra-se um elemento do vetor w.
10 20 30 40 50 60 70 80 90
−2
−
1.5
−1
−
0.5
0
0.5
1
1.5
2
Figura 4.8 - Representação gráfica de w1 x freqüência
10 20 30 40 50 60 70 80 90
−2
−1.8
−1.6
−1.4
−1.2
−1
−0.8
−0.6
−0.4
−0.2
0

117
Figura 4.9 - Representação gráfica de w2 x freqüência
Portanto, verifica-se que w(2) é uma constante para qualquer freqüência, e que
w(1) possui valores cíclicos. A solução ótima do vetor w permite minimizar diretamente
o valor do erro quadrático médio do sinal desejado.
4.4.5 Solução subótima usando aproximação estocástica do
Gradiente
O padrão para utilizar um filtro transversal é encontrar uma melhor solução para
seu vetor de coeficientes que satisfaz as equações de Wiener. Uma maneira de fazer isto
seria resolver as equações do sistema por algum método analítico. Este procedimento é
bastante trabalhoso, apresentando sérias dificuldades computacionais, especialmente
para filtros de ordem elevada, ou quando a quantidade de dados de entrada for elevada.
Um procedimento alternativo é o “método de descida do gradiente”, muito usado em
otimização. Logo, basta encontrar o mínimo valor para o erro quadrático médio,
baseado nas equações de Wiener.
Seja
(
)
(n)ξ
∇
, o vetor de gradiente no tempo n e a(n) o vetor de coeficientes no
tempo n. De acordo com o método da descida do gradiente, a atualização do vetor de
coeficientes no tempo n+1 é feita usando a relação recursiva abaixo:
()
](n)[
2
1
(n)1)(n ξaa −∇+=+
µ
(4.35)
onde, µ é uma constante de valor positivo real, denominada “passo de adaptação”. O
fator ½ é usado apenas por conveniência (HAYKIN, 1991).
O vetor gradiente
(
)
(n)ξ∇ é representado por:
(
)
(n)ξ∇ =
⎥
⎥
⎥
⎥
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎢
⎢
⎢
⎢
⎣
⎡
∂
∂
∂
∂
∂
∂
−
)(
)(
....
)(
)(
)(
)(
1
1
0
na
n
na
n
na
n
N
ξ
ξ
ξ

118
= -2
p + 2 R a(n) (4.36)
onde, na primeira linha, )(/)( nan
k
∂
∂
ξ
é a derivada parcial da função custo )(n
ξ
com
relação à
. Para a aplicação do algoritmo de descida do gradiente, assumimos a
correlação da matriz
R e a correlação cruzada do vetor rp que são conhecidos quando
calculamos o vetor gradiente
)(na
k
(
)
(n)ξ
∇
para os valores do vetor de coeficientes a(n).
Então, substituindo (4.36) em (4.35), calcular-se o vetor de coeficientes
a(n+1)
usando a relação recursiva abaixo:
](n)[(n)1)(n
Rarpaa
−
+
=+
µ
n= 0,1,2,....
(4.37)
O parâmetro µ controla o tamanho da aplicação da correção, incrementando o
vetor de coeficientes no processo iterativo a cada ciclo do algoritmo.
Com isso, o cálculo dos coeficientes na Predição Linear Direta é feito a partir
das análises anteriores, pelo método da descida do gradiente. A solução normalmente
adotada consiste em aproximar os cálculos de forma a não comprometer os resultados
estatísticos, diminuindo o tempo de processamento computacional, através da seguinte
simplificação:
)1(.(n).2 −−≅
∂
knxe
a
p
k
ξ
(4.38)
Deve-se observar que, do ponto de vista da Estatística formal, a aproximação
(4.38) pode parecer incoerente e muito grosseira. Todavia, a prática do processamento
de sinais, desde a década de 60 (HAYKIN, 2000), revela que a equação (4.38) se mostra
eficiente para as mais variadas aplicações.
Consequentemente, a atualização dos novos coeficientes a partir de coeficientes
anteriores, após calculado o erro de predição, utiliza a equação:
(n).(n).2(n)1)(n
xaa
p
e
µ
−
=
+
(4.39)
Essa solução normalmente adotada pela literatura consiste em aproximar os
cálculos de forma a não comprometer os resultados estatísticos, diminuindo o tempo de
processamento computacional.
119
4.5 Predição Linear Reversa (“backward”)
Na seção anterior, discutiu-se como uma amostra x(n) pode ser estimada a partir
de N amostras precedentes {x(n-1), x(n-2), ..., x(n-N+1)}. O objetivo desta seção
consiste em fazer justamente o contrário: estimar a amostra x(n-N) a partir das N
amostras posteriores.
4.5.1 Modelo Auto-Regressivo (AR) Reverso
Modelamos um sinal a partir de parâmetros estabelecidos. A técnica de predição
linear para este modelo, considera uma relação linear, conforme abaixo:
)1(...)()()(
110
+
−
+
+
+
=−
−
NnvgnvgnvgNnx
n
(4.40)
∑
−
=
−=−
1
0
)()(
N
i
i
invgNnx
(4.41)
onde
{
são coeficientes (constantes) e N é a ordem do modelo.
}
110
,...,,
−N
ggg
4.5.2 Modelo da Predição Linear Reversa
Este modelo prevê uma amostra de sinal a partir de suas amostras analisadas
posteriormente.
∑
=
+−=−
N
k
k
knxgNnx
1
^
)1()(
(4.42)
)1(.....)1()()(
21
^
+−++−+=− NnxgnxgnxgNnx
N
(4.43)
onde o valor de x previsto
é calculado a partir de valores já
conhecidos.{x(n),....,x(n-N+1)}.
⎟
⎠
⎞
⎜
⎝
⎛
− )(
^
Nnx
Para calcularmos o erro de predição, fazemos a diferença entre o sinal real e o
sinal predito. Ou seja,
120
)()(
^
NnxNnxe
pr
−−−=
(4.44)
onde
é o erro de predição e (n-N) é calculado a partir de : {x(n), x(n-1),...,x(n-
N+1)}.
pr
e
^
x
4.5.3 Critérios para uma boa predição
Analisaremos os dois critérios citados na seção 4.4.3, para saber se o modelo
está funcionando adequadamente.
1 -
→0
pr
e
2 -
→0
{}
)(
2
^
neE
pr
Os dois critérios devem atingir o menor valor possível, para que esse modelo
seja perfeito para o sinal analisado.
4.5.4 Solução Ótima de Wiener para Predição Linear Reversa
Para se resolver as equações anteriores a partir das Equações de Wiener-Hopf,
têm-se três características importantes, discutidas para a Predição Linear Direta. Agora,
analisaremos as mesmas para a Predição Linear Reversa.
1) Seja
x(n) o vetor de ordem 1 x N que contém os dados de entrada:
T
Nnxnxnx ])1(...)1()([(n) +−−=x
(4.45)
A matriz de correlação dos valores de entrada de ordem N por N será:
](n)(n)[
T
xxR E=
(4.46)
A forma estendida da matriz de correlação
R é dada em (4.25).
2) O vetor de correlação cruzada entre os valores de entrada {x(n), x(n-1),...,x(n-N+1)}
e a saída desejada x(n-N) é:
121
)]((n)[ NnxE
−
=
xr
pr
=
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎣
⎡
−
)1(
...
)1(
)(
r
Nr
nr
(4.47)
A forma estendida dessa correlação também está descrita em (4.26).
3) A variância da saída desejada x(n-N) é igual a r(0).
Pode-se analisar a predição linear reversa e, consequentemente, adaptar as
Equações de Wiener (4.4)-(4.6) para resolver o problema para entradas estacionárias:
Rg =
pr
r
(4.48)
onde
[
]
N
ggg ..
21
=g são os coeficientes do filtro.
Similarmente, usando (4.14) em (4.48), produzimos a seguinte expressão para o
erro de predição reversa:
gr
pr
T
pr
re −= )0(
(4.49)
Aqui, novamente, tem-se um vetor de coeficientes
g de ordem N por 1 para
predição linear reversa, e
é o erro de predição reversa, definidos por um conjunto de
valores da função de autocorrelação no intervalo {0,1,...., N}.
pr
e
4.5.5 Solução subótima usando aproximação estocástica do
Gradiente para a Predição Reversa
Adapta-se o método da descida do gradiente, descrito em 4.4.5, para a Predição
Linear Reversa. De acordo com (4.35) e (4.36), calcula-se o vetor de coeficientes
g(n+1), usando a relação recursiva abaixo:
](n)[(n)1)(n
Rgrgg
pr
−
+
=+
µ
n=0, 1, 2, ...
(4.50)
Em seguida, atualizam-se os novos coeficientes a partir de coeficientes
anteriores, depois de se calcular o erro de predição reversa, de acordo com:
122
(n)).(.2.(n)1)(n
xgg ne
pr
µ
−
=
+
(4.51)
onde, µ é o passo de adaptação.
Uma forma interessante de abordar a predição reversa consiste em ver o preditor
regressivo atuando nas N amostras armazenadas, de forma a gerar uma estimativa da
amostra que acabou de sair do filtro. Nesse contexto, a predição reversa faz tanto (ou
tão pouco) sentido quanto à predição direta, pois ambas procuram estimar algo que já é
conhecido. De fato, em nosso caso, não estamos interessados em estimativas geradas
pelos preditores, mas sim nos coeficientes e na ordem do filtro de predição.
4.6 Estatísticas de Ordem Elevada (EOE)
A teoria de EOE é uma área que desenvolve a análise do sinal com aplicações na
ciência e na engenharia, em particular em telecomunicações, radar, sonar, geologia,
processamento de imagem, processamento de voz, biomedicina, oceanografia, física,
economia e mecânica dos fluidos (NIKIAS, 1993).
O interesse na utilização de EOE reside na possibilidade de refinar a
caracterização da função de densidade de probabilidade conjunta de um processo
estocástico; bem como nas propriedades dos cumulantes e na biunivocidade de
representação entre os espectros de ordem elevada (bi e triespectro, por exemplo) e o
processo estocástico sob análise. Esta última razão permite resolver as limitações da
análise baseada na densidade espectral de potência, a qual não inclui informação de fase
a respeito do sinal.
As vantagens de técnicas baseadas em EOE, descritas no parágrafo anterior,
contrastam com duas limitações principais (PAPOULIS, 1990). Primeiramente, o custo
computacional dos algoritmos é muito elevado. Em seguida, a convergência dos
estimadores estatísticos é demorada, exigindo, em geral, uma grande quantidade de
amostra do sinal.
Consequentemente existem algumas razões para se utilizar a EOE, tais como
(CADZOW, 1996): processar um sinal para extrair as informações devido aos desvios
da gaussianidade através do biespectro e triespectro; além da determinação da não-

123
linearidade de uma série temporal através do teste de Hinich (PAPOULIS, 1990). As
EOE também transformam o sinal com ruído, permitindo sua detecção e reconstrução
precisa.
4.6.1 Funções com uma variável aleatória (CADZOW, 1996)
A função característica, associada à variável aleatória x é definida como:
{}
xjxj
xx
eEdxexf
ωω
ωφ
==
∫
∞
∞−
)()(
(4.52)
Onde
representa o operador esperança matemática.
{}
•E
A função )(
ω
φ
x
corresponde ao complexo conjugado da Transformada de
Fourier da função de densidade de probabilidade, sendo também denominada função
geradora dos momentos. Assim, esta possui propriedades associadas à transformada de
Fourier. Logo, a transformada inversa é definida como:
ωωφ
π
ω
dexf
xj
xx
∫
∞
∞−
−
= )(
2
1
)(
(4.53)
A função geradora dos momentos possui as seguintes propriedades:
)()(
ωφωφ
ω
x
aj
y
eaxy =⇔+=
(4.54a)
)()(
ω
φ
ω
φ
aaxy
xy
=
⇔
=
(4.54b)
)()()(
2121
ω
φ
ω
φ
ω
φ
xxy
xxy
=
⇔
+=
(4.54c)
1)0(
=
x
φ
(4.54d)
ωωφ
todopara
x
1)( ≤
(4.54e)
)()(
*
ωφωφ
−=
xx
(4.54f)
Onde a é um escalar nas duas primeiras propriedades; x
1
e x
2
são variáveis aleatórias
independentes, na terceira propriedade.

124
Se o momento de ordem m da variável aleatória x existir, é possível fazer uma
expansão da série de Taylor, associando os momentos à função característica, como
mostrado logo abaixo:
{}
)()(
!
1
)(
0
ωωωφ
m
kk
m
k
x
ojxE
k
+=
∑
=
(4.55)
{}
mkparaxEj
d
d
kk
k
x
k
,...,3,2,1)(
)(
0
==
=
ω
ω
ωφ
(4.56)
Onde )(
ω
m
o representa a função resto.
A função geradora dos cumulantes, ou segunda função característica, é
especificada logo abaixo:
[
]
}]{ln[)(ln)(
xj
xx
eE
ω
ωφω
ψ
==
(4.57)
Analogamente às propriedades da função geradora dos momentos (4.54a) –
(4.54f), definem-se as seguintes propriedades para a função geradora dos cumulantes.
)()(
ω
ψ
ω
ω
ψ
xy
ajaxy
+
=
⇔
+=
(4.58a)
)()(
ω
ψ
ω
ψ
aaxy
xy
=
⇔
=
(4.58b)
)()()(
2121
ω
ψ
ω
ψ
ω
ψ
xxy
xxy
+
=
⇔
+=
(4.58c)
Onde a é um escalar nas duas primeiras propriedades; x
1
e x
2
são variáveis aleatórias
independentes, na terceira propriedade.
O cumulante da variável aleatória x é definido por:
,...3,2,1
)(
)()(
0
=−=
=
kpara
d
d
jkC
k
x
k
k
x
ω
ω
ωψ
(4.59)
Portanto, se a variável aleatória x for um valor real, mostra-se que estes
cumulantes são valores reais. A partir de (4.59), )(
ω
ψ
x
pode ser aproximada por:
)())((
!
1
)(
1
ωωω
ψ
m
k
x
m
k
x
ojkC
k
+=
∑
=
(4.60)
Onde )(
ω
m
o é a função resto.

125
4.6.2 Funções com várias variáveis aleatórias (NIKIAS, 1993)
Dado o conjunto de m variáveis reais randômicas { , os momentos
de ordem
}..,,,
21 m
xxx
m
kkkkr
+
+++= ...
321
, onde é um inteiro, são definidos por (PAPOULIS,
1990):
i
k
Mom = [
]
km
m
kk
xxx ...,,,
2
2
1
1
∆
{
}
=
km
m
kk
xxxE ...
2
2
1
1
0...
2
2
1
1
21
21
...
)...,,,(
)(
====
∂∂∂
Φ∂
−
m
km
m
kk
m
r
r
j
ωωω
ωωω
ωωω
(4.61)
))}...(exp({)...,,,(
221121 mmm
xxxjE
ω
ω
ω
ω
ω
ω
+
+
+
∆Φ
(4.62)
Onde )...,,,(
21 m
ω
ω
ω
Φ representa a primeira função característica.
A segunda função característica de um conjunto de variáveis aleatórias
multidimensionais é definida logo abaixo (PAPOULIS, 1990):
)]...,,,([ln)...,,,(
~
2121 mm
ωωωωωω
Φ∆Ψ
(4.63)
A função cumulante (também denominada “semi-invariante”) de ordem r, do
mesmo conjunto de variáveis randômicas, é definida como o coeficiente da expansão da
série de Taylor da segunda função característica em torno de zero; isto é:
]...,,,[
2
2
1
1
km
m
kk
xxxCum ∆
0...
2
2
1
1
21
21
...
)...,,,(
~
)(
====
∂∂∂
Ψ∂
−
m
km
m
kk
m
r
r
j
ωωω
ωωω
ωωω
(4.64)
As relações entre momentos e cumulantes podem ser expressas pelas equações
logo abaixo:
M
1
= Mom[x
1
] = E{ }
1
x
(4.65a)
M
2
= Mom[x
1,
x
1
] = E{ }
2
1
x
(4.65b)
M
3
= Mom[x
1,
x
1,
x
1
] = E{ }
3
1
x
(4.65c)
M
4
= Mom[x
1,
x
1,
x
1,
x
1
] = E{ }
4
1
x
(4.65d)
111
][ MxCumC
=
=
(4.65e)

126
2
12112
],[ MMxxCumC −==
(4.65f)
3
11231113
23],,[ MMMMxxxCumC +−==
(4.65g)
4
1
2
12
2
213411114
61234],,,[ MMMMMMMxxxxCumC −+−−==
(4.65h)
Se E{
} = M
1
x
1
= 0, então
2
2443322
3,, MMCMCMC −===
.
No caso específico de um processo estocástico, em função da variável tempo, se
as m variáveis aleatórias
{}
)(...,),(),(
121 −m
xxx
τ
τ
τ
forem estacionárias, pode-se definir
as multicorrelações de ordem elevada como segue:
{
}
)(..)().()...,,,(
121121 −−
−
−
−
∆
mmmx
xxxEM
τ
τ
τ
τ
τ
τ
"
(4.66)
Exemplo 4.2:
Considere as três funções de densidade de probabilidade (pdfs)
simétricas dos tipos Laplaciana, Gaussiana e Uniforme. Os momentos de M
m
de ordem
m = 1,2,3,4 podem ser gerados de (PAPOULIS, 1990):
dxxfxM
m
m
)(
∫
∞
∞−
=
(4.67)
onde f(x) é a função de densidade de probabilidade (pdf). Assim, calcula-se a função
característica, definida por:
∫
∞
∞−
=Φ dxxjxf
x
)(exp)()(
ωω
(4.68)
Os cumulantes
, m = 1,2,3,4 são facilmente calculados através dos momentos
em (4.65a)-(4.65h).
m
C
As Figuras 4.10-4.12 e as Tabelas 4.2-4.4 ilustram os momentos e os cumulantes
de ordem 1 a 4 das pdfs citadas logo acima. Os gráficos foram gerados em ambiente
MATLAB, considerando a variável aleatória x.
(a) Função Laplaciana: f(x) = 0.5 exp (-|x|)

127
−6 −4 −2 0 2 4 6
0
0
.05
0.1
0
.15
0.2
0
.25
0.3
0
.35
0.4
0
.45
0.5
Tabela 4.2 – Função Laplaciana
m M
m
C
m
1 0 0
2 2 2
3 0 0
4 24 12
Figura 4.10 – Gráfico da Função Laplaciana, com seus momentos e cumulantes de ordem 1 a 4.
(b) Função Gaussiana: (x) = 0.5 exp
2
2
2
.2/
2
σπ
σ
⎭
⎬
⎫
⎩
⎨
⎧
−
x
, onde 88.2
2
=
σ
−5 −4 −3 −2 −1 0 1 2 3 4 5
0
.02
0
.04
0
.06
0
.08
0.1
0
.12
0
.14
Tabela 4.3 – Função Gaussiana
m M
m
C
m
1 0 0
2
2
σ
2
σ
3 0 0
4
3
4
σ
0
Figura 4.11 – Gráfico da Função Gaussiana, com seus momentos e cumulantes de ordem 1 a 4.
(c) Função Uniforme: f(x) =
c2
1
, onde c = -0.5 a 0.5
−5 −4 −3 −2 −1 0 1 2 3 4 5
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Tabela 4.4 – Função Uniforme
m M
m
C
m
1 0 0
2 c
2
/3 c
2
/3
3 0 0
4 c
4
/5 -2c
4
/15
Figura 4.12 – Gráfico da Função Uniforme, com seus momentos e cumulantes de 1 a 4 ordem.

128
Portanto, verifica-se que, como todas as funções pdfs analisadas são simétricas,
todos M
m
e C
m
ímpares apresentam valores zero. Para o exemplo da Gaussiana (Figura
4.11 e Tabela 4.3), todos os seus cumulantes
C
m
de ordem maior que a segunda (n > 2)
também são zero.
4.6.3 O ruído branco e os índices de descorrelação
Um “ruído branco puro de ordem elevada de ordem p”, ou sinal pw(n), é
definido pelas seguintes equações ((NIKIAS, 1993), (BONDON, 1991)):
p2;
);,...,,(pwR),...,,pw(C
121121m
<<∈
=
∗
−−
mm
mmm
τ
τ
τ
δ
τ
τ
τ
(4.69)
Onde:
),...,,pw(C
121m −m
τ
τ
τ
: cumulante de ordem m de pw(n);
∈
i
τ
∗
: conjunto de números naturais não-nulos
pwR
m
: constante real
),...,,(
121 −m
τ
τ
τ
δ
: Símbolo de Kronecker generalizado
⎩
⎨
⎧
====
∆=
−
−
contráriodo
se
m
m
0
0...;1
),,(
121
121
τττ
τττδ
(4.70)
Os cumulantes de ordem 2, 3, 4 do processo estocástico x(n) são
respectivamente estimados como a seguir (NIKIAS, 1993), supondo x(n) estacionário:
∑
−
=
−≤≤+
−
=
1
1
2
^
0);().(
1
1
)(
K
i
x
iKixix
K
C
τττ
(4.71)
iKiK
ixixix
K
C
K
i
x
−≤≤−≤≤
++
−
=
∑
−
=
21
2
1
2121
3
^
0;0
;)().().(
2
1
),(
ττ
ττττ
(4.72)
iKiKiK
ixixixix
K
CC
K
i
xx
−≤≤−≤≤−≤≤
+++
−
+−=
∑
−
=
321
3
1
321
2
21
2
^
321
4
^
0;0;0
)().().().(
3
1
)),(.(3),,(
τττ
ττττττττ
(4.73)

129
Onde K é o comprimento da série temporal, ou o número total das amostras de x(n)
disponíveis para os cálculos.
Os Índices de Descorrelação Elevada (IDE) W2, W3, W4 de um processo
estocástico geral x(n) são definidos abaixo, e discutidos detalhadamente em ((NIKIAS,
1993), (DESTRO-FILHO, 2002), (DESTRO-FILHO, 2003)):
4,3,2;)(
2
== ipi WW
(4.74)
4,3,2;
,...)0,0,0(C
),...,,(C.....,...)0,0,0(C
pX
121pX
12 1
pX
1
=
−
=
−
== =
∑∑ ∑
−
pW
p
qq q
p
p
τττ
ττ τ
(4.75)
Os valores de q, usados nas equações (4.74)-(4.75), dependem do sinal a ser
considerado. Particularmente, q não deve ser maior que o “intervalo de
estacionariedade”, isto é, o período de tempo durante o qual o sinal mantém suas
características estatísticas constantes.
Deve-se observar que, de acordo com (4.69)-(4.70),
atinge seu valor máximo
no caso do “ruído branco puro de ordem elevada”, para o qual Wm = 1 para todo valor
de m dentro do intervalo 2 < m < p. Observe também que, baseado em resultados
teóricos ((NIKIAS, 1993), (BONDON, 1991)), se o processo estocástico for periódico
(por exemplo, neurônios em potenciação), então Wm apresenta valores muito baixos,
incluindo amplitudes com valores próximos de zero. Relativamente à gaussianidade, o
processo estocástico gaussiano apresenta cumulantes de 3ª e 4ª ordem próximos à zero,
conduzindo a IDE W2, W3, W4 de amplitudes muito elevadas, sempre maiores que 1
((NIKIAS, 1993), (BONDON, 1991)).
p
W
Em resumo, o IDE permite avaliar características estatísticas ligadas à
periodicidade, gaussianidade e descorrelação de um processo estocástico qualquer.
4.7 Conclusão
Neste capítulo analisaram-se as ferramentas simples, em conformidade com os
desafios estabelecidos no Capítulo 2, para lidar com a não-estacionariedade e a não-

130
linearidade de sinais biológicos, particularmente os sinais de MEA e EEG. Quando se
trata um sinal não-estacionário, aplica-se a filtragem adaptativa. Já para lidar com um
sinal resultante de um sistema não-linear, aplica-se a teoria de EOE para refinar a
caracterização estatística do sinal em questão.
O registro de sinais bioelétricos é de grande importância, porém durante a
aquisição, estes sinais encontram-se invariavelmente associados a um grande número de
interferências das mais diversas origens. Uma das possíveis técnicas para a eliminação
dessas interferências é o uso de filtros adaptativos. Estes são ferramentas extremamente
úteis na área de processamento de sinais, principalmente em aplicações envolvendo
tempo real, onde pouca ou quase nenhuma informação sobre a estrutura estatística dos
sinais analisados é conhecida a priori (WIDROW, 1985).
Dentre as diversas técnicas de filtragem adaptativa, destaca-se a predição linear,
já estabelecida desde 1960 (WIDROW, 1985), permitindo alcançar bom desempenho
através de algoritmos de baixa complexidade computacional. Na Tabela 4.5 abaixo,
apresenta-se um pequeno resumo das características associadas aos filtros de Wiener, à
Predição Linear Direta e à Predição Linear Reversa.
Tabela 4.5 - Sumário das variáveis do filtro de Wiener (adaptada de (HAYKIN, 1991))
Quantidade Filtro de Wiener Predição Linear
Direta
Predição Linear
Reversa
Vetor de entrada
x(n) x(n-1) x(n)
Saída desejada
d(n) x (n) x(n-N)
Vetor de coeficientes
W A G
Estimativa de erro
e(n)
)(ne
p
pr
e (n)
Matriz de correlação
R R R
Vetor de correlação
cruzada
P Rp
pr
r
Erro médio
quadrático
£
{
}
)(
2
^
neE
p
{}
)(
2
^
neE
pr
Desta maneira, a predição linear só tem sentido quando o processo estocástico é
suposto estacionário, pois do contrário, a matriz
R e o vetor P não são univocamente
131
definidos, mesmo para N suficientemente grande. Ou seja, se a série temporal resulta de
um processo estocástico não-estacionário,
R e P variam ao longo da série, invalidando o
uso de (4.14) para a obtenção do vetor de coeficientes
W. A solução algumas vezes
adotada é assumir que a série temporal seja estacionária em intervalos, adaptando-se
R e
P para cada intervalo.
O processamento de sinais baseado em estatística de ordem elevada (NIKIAS,
1993) apresenta características muito interessantes para aplicações em engenharia
neural. Por exemplo, os efeitos de ruído gaussiano, que perturbam freqüentemente as
medidas neurofisiológicas, podem ser minimizados a partir da seguinte propriedade. Os
cumulantes de um sinal composto por atividade neural espontânea (ANE) e o ruído
gaussiano são exatamente os mesmos que os cumulantes associados ao sinal ANE puro.
Além disso, as correlações de ordem elevada fornecem uma informação mais
precisa da função de densidade de probabilidade, permitindo assim uma classificação e
uma análise mais exatas do sinal, em aplicações onde a relação sinal/ruído (S/R) é muito
baixa. Em (NIKIAS, 1993), os autores apresentam exemplos de diversos algoritmos de
classificação, baseados em ordens elevadas, que resultam em desempenho muito
eficiente, para aplicações com relações S/R abaixo de 5 dB. Por outro lado, as
limitações da teoria de EOE não devem ser esquecidas, levando em conta o alto custo
computacional dos algoritmos e a lentidão de convergência dos estimadores.
No próximo capítulo, as ferramentas aqui discutidas são aplicadas à análise de
sinais MEA e EEG.
132
Capítulo 5
Simulações e Resultados
5.1 Introdução
O objetivo deste capítulo é estabelecer modelos AR simples para os sinais
biológicos, medidos através do EEG e das MEAs. Além disso, investigou-se a estrutura
estatística do sinal resultante da MEA em termos do conceito de ruído branco.
Em particular, a análise do sinal EEG contendo crises epilépticas exige a
estimação do tempo de estacionariedade, que corresponde ao intervalo de tempo durante
o qual as características estatísticas do sinal se mantêm constantes (BLESIC, 2003). É
importante frisar que o tempo de estacionariedade (TE) associado a trechos de EEG
contendo crises epilépticas estabelece um limitante máximo para a previsibilidade da
ocorrência de crises, conforme discutido no Capítulo 3.
De fato, o tempo de estacionariedade pode ser interpretado como o tempo médio
para que uma mudança fisiológica ocorra no tecido considerado, de tal forma a alterar a
resposta elétrica das células envolvidas e, conseqüentemente, a informação associada à
atividade elétrica neural (SHOUVAL, 2005). Sendo assim, estabelecendo-se um
conhecimento sobre as características do sinal EEG, é possível buscar soluções para as
patologias que envolvem a alteração da neurodinâmica, associada à transmissão de
informações através do Sistema Nervoso Central. A mesma observação pode ser feita
133
para o caso do sinal MEA, onde as alterações estatísticas poderiam ser correlacionadas à
alterações da conectividade entre os neurônios em cultura.
Inicialmente, descreve-se a aquisição dos sinais, os quais foram utilizados
durante as simulações. Depois, discutem-se os critérios de desempenho do filtro de
predição linear, em particular os tempos de convergência e de estacionariedade, que
permitem avaliar o desempenho da filtragem linear adaptativa em um neuroimplante.
Além disso, os limitantes práticos dos índices de descorrelação são estabelecidos,
permitindo agrupar canais da MEA com comportamento estatístico semelhante.
5.2 Aquisição dos Sinais
O conjunto de sinais descrito a seguir foi medido por outros grupos de pesquisa,
que registraram os dados sob formato digital.
(A) Medidas in vivo dos neurônios dos invertebrados
A seqüência temporal de registros intracelulares da atividade espontânea dos
neurônios do gânglio cerebral do caramujo Tritonia diomedea foi obtido in vivo,
conforme descrito em (PEIXOTO, 2002). O animal foi cortado aberto, o tecido
conjuntivo extraído e o cérebro exposto. Sem dissecção, mas mantendo o cérebro em
contato com a água do mar, um eletrodo de vidro, usado para a gravação intracelular, foi
introduzido dentro das células identificadas. Os sinais foram registrados
ininterruptamente durante pelo menos seis horas. O tempo de amostragem utilizado foi
de 0,1 milissegundos.
Um exemplo do sinal da atividade espontânea é apresentado na Figura 5.1. Estes
sinais serão denominados i(n), onde o número natural “n” denota o tempo discreto.
Observe que i(n) não é uma série temporal de intervalos entre “spikes”, mas sim as
amplitudes reais do sinal de atividade neural espontânea (ANE).

134
Figura 5.1 – Potenciais de ação associados a neurônios do gânglio cerebral do
caramujo Tritonia diomedea (adaptada de (PEIXOTO, 2002))
(B) Sinal de Matriz Multieletrodo (MEA)
Os sinais serão denominados m(n), consistindo nas amplitudes de um sinal ANE,
medidos em culturas primárias de neurônios corticais de galinha, cultivados no meio
Neuro Basal. As medidas ocorreram por meio de MEAs contendo sessenta (60)
microeletrodos planares (MEA, Sistema Multicanal), após 20 dias in-vitro, a fim de
permitir a maturação das sinapses entre os neurônios. O tempo de amostragem é de
quatro milessegundos (4 ms). Detalhes adicionais são apresentados em (NOVELLINO,
2003).
(C) Sinal Eletroencefalograma (EEG)
Esta documentação foi feita a partir do banco de dados fornecidos pela
Universidade de Tampere (Finlândia), detalhados em (BERNARDI, 1999), obtidos
através de (URL 11). Este banco de dados é constituído por 9 arquivos, onde em cada
um existe a presença de crises epilépticas do tipo lobotemporal, no formato:
tv00XXX0.rec, onde:
tv0001B0.REC e tv0004B0.REC ⇒ contém pequenas descargas epilépticas
de um tumor.
tv0005B0.REC e tv0008.REC ⇒ contém tipo irregular de descarga
epiléptica.
tv0003A0.REC ⇒ contém três descargas epilépticas.
tv0009B0.REC e tv0012B0.REC ⇒ contém crises epilépticas com artefato
eletromiográfico sobre o EEG.
135
tv0013B0.REC e tv0014B0.REC ⇒ contém crises epilépticas psicomotoras.
Cada arquivo destes contém o sinal proveniente de 4 eletrodos, arranjados no
formato F8-C4, F7-C3, T6-C2 e T5-C1, medidos durante 160 segundos. Cada arquivo
foi subdividido em duas pastas, denominadas NãoCrise e Crise, onde a primeira inclui
trechos de sinal que não contém indícios evidentes de crises epilépticas; e a segunda
inclui trechos que apresentam algum tipo de crise epiléptica.
Os dados foram coletados na superfície do escalpo com eletrodos bipolares
compostos por uma liga Ag-AgCl, com taxa de amostragem de 200 Hz. Foi aplicado um
filtro passa-baixa em 70 Hz. Os eletrodos são fixados no escalpo do paciente
obedecendo ao sistema internacional 10-20 (SAAB, 2005), possuindo no total 21
eletrodos: dois fixados nas orelhas e o restante nas regiões do escalpo. Os dados foram
adquiridos de três homens e duas mulheres, com idade média de 28.87± 15.27.
5.3 Ferramentas e Metodologia
O computador utilizado para o processamento dos dados possui as seguintes
características:
Processador AMD Sempron 2200, relógio 1.49GHz;
Memória RAM de 1 G Bytes;
Memória ROM de 512 M Bytes.
5.3.1 Processamento dos dados e critérios utilizados na Predição
Linear
As equações da seção 4.4 foram simuladas através do seguinte procedimento
iterativo:
(início); 1=n
Determina-se, de forma aleatória pela função rand(n), valores para os
coeficientes iniciais do filtro:

136
()
[
]
T
n 0.3 0 0.9...7.011 a ==
(5.1)
Para até L1+n
MAX
; onde L
MAX
= 2000 (sinal EEG) ou L
MAX
= 3000 (sinal
MEA);
No instante de tempo n, deve-se conhecer o valor da amostra atual e das N
amostras anteriores a esta:
{
,
()
nx
()
1
−
nx ,
(
)
2
−
nx , ...,
(
)
Nnx
−
}
(5.2)
Calcula-se a amostra atual (
(
)
nx
ˆ
) usando a equação de predição (4.22);
)(...)2()1()(
110
^
Nnxanxanxanx
N
−++−+−≅
−
(5.3)
Calcula-se o erro de predição (
(
)
ne
p
) usando (4.23);
)()()(
^
nxnxne
p
−=
(5.4)
Calculam-se os novos coeficientes a partir dos coeficientes anteriores, dos
valores das amostras e do erro de predição, através da equação (4.39);
(n)).(.2(n)1)(n
xaa ne
p
µ
−
=
+
(5.5)
Calcula-se de uma estimativa do erro quadrado médio (eqm) a partir dos valores
do erro de predição:
()
}{
()
∑
=
≅
n
j
pp
je
n
neE
1
22
1
ˆ
(5.6)
As equações
(5.1)-(5.6) foram implementadas em ambiente MATLAB.
Os resultados apresentados no item seguinte são gráficos cuja escala horizontal
corresponde ao tempo discreto n, expresso em quantidade de iterações (vide Figura 5.2).
A escala vertical corresponde ao erro quadrático médio instantâneo, definido em (5.6).

137
Figura 5.2 - Exemplo de um gráfico típico de eqm x n. Eixo x: tempo
Eixo y: erro quadrático médio
A partir desses gráficos, foram estimadas as seguintes grandezas, que definem
critérios de desempenho para o funcionamento do filtro de predição, ou seja, expressam
a eficiência do modelo do sinal construído com base na técnica de predição linear.
Erro Quadrado Médio de Regime Permanente (EQMr): refere-se ao valor do
erro quadrado médio após a convergência do filtro para uma situação estável.
Aqui, “regime permanente” significa o instante de tempo T a partir do qual o
gráfico eqm x n apresenta amplitudes de eqm com baixa oscilação, onde a
diferença da amplitude máxima não ultrapassa 10% do valor da amplitude
mínima, conforme ilustrado na
Figura 5.3. O valor de EQMr é estabelecido
então como o valor médio do EQMr considerando as últimas 200 amostras, logo
após a convergência. Desta forma, a amplitude de EQMr expressa a precisão da
predição realizada pelo filtro.

138
Figura 5.3 - Exemplo de EQMr, em um gráfico típico de eqm x n. O valor de EQMr,
calculado como média entre n=400 e n=600, é
.
68≅EQMr
Pico Máximo (PM): refere-se ao máximo valor de erro quadrado médio (eqm),
que ocorre logo após o início do processo de filtragem. Esse pico caracteriza a
estabilidade do filtro, visto que, para situações de não-convergência, sua
amplitude é excessiva. Na Figura 5.4
, o PM corresponde à amplitude máxima do
gráfico eqm x n, que ocorre para
600
≅
n .
Figura 5.4 - Exemplo de um gráfico típico de eqm x n em que o PM representa o
ponto mais alto do gráfico, ocorrendo em n ≈ 600.

139
Tempo de Convergência (TC): refere-se ao tempo gasto, em iterações, para que
o gráfico de eqm x iterações apresente variações de amplitude confinadas no
intervalo
]
[
EQMrxEQMrx 1,1;99,0 . Dessa forma, TC caracteriza a capacidade
do filtro em responder em tempo real ao sinal de entrada a ser modelado. Na
Figura 5.5, o TC está representado pelo encontro do eqm e a reta que representa
o valor EQMr + 10%. Assim sendo, 600
≅
TC .
Figura 5.5 – Exemplo de um gráfico típico de eqm x n em que o TC representa a
convergência do sinal MEA TC
≅
600.
Tempo de Estacionariedade (TE): refere-se ao tempo durante o qual as
características estatísticas do sinal modelado se mantêm inalteradas. Este tempo
é determinado pelo primeiro instante de tempo, logo após TC, em que a
amplitude do gráfico eqm x n atinge o valor 1.7 x EQMr. Na Figura 5.6,
observa-se um pico inicial do eqm, que decai até atingir o regime permanente no
tempo n = 400. Todavia, a partir do tempo n = 500, observa-se um aumento
abrupto do eqm, de tal forma que em n = 600 o eqm atinge o valor 1.7 x EQMr.
Este aumento abrupto revela alterações profundas na estrutura estatística do
sinal, donde TE
600. ≅

140
Figura 5.6 - Exemplo de gráfico típico de eqm x n para o caso de um sinal EEG com crise TE 600
≅
.
Cumpre destacar que o objetivo final do trabalho computacional consiste em
realizar uma análise dos gráficos de erro quadrado médio (eqm) versus número de
iterações (n), tendo por objetivo encontrar os parâmetros de ordem do modelo (
) e do
passo de adaptação (
N
µ
) corretos, para permitir que um compromisso ótimo envolvendo
estabilidade do modelo (que se traduz por mínimo PM), velocidade de resposta do filtro
em tempo real (que se traduz por mínimo tempo de convergência TC) e precisão de
previsão (mínimo erro quadrado médio de regime permanente EQMr) possa ser
alcançado.
5.3.2 Processamento de dados utilizado para estimação dos índices de
descorrelação
Consideraram-se cinco tipos de sinais nos cálculos.
O primeiro é o i(n) (vide Figura 5.1), ou sinal da atividade neural espontânea do
caramujo.
O segundo é m(n), ou o conjunto de registros de 60 canais MEA (veja o item 5.2
B).
O terceiro sinal é um ruído branco de ordem elevada (NIKIAS, 1993), que será
identificado aqui como hw(n). Ele é obtido por meio de um gerador especial de

141
números aleatórios, que fornece uma série temporal com as correlações de
ordem elevada insignificantes, até a 4ª ordem.
O quarto é um sinal cíclico de clock, que é identificado como o c(n). Esta série
periódica é usada atualmente para a sincronização de Sistemas de
Posicionamento Global (GPS), e foi gerado por um software de acordo com um
procedimento definido em (PARKINSON, 1995).
O quinto é uma série temporal identificada como g(n), e foi obtida por meio de
um gerador de números aleatórios padrão, para fornecer um processo estocástico
correlacionado gaussiano.
Para cada sinal {i(n), m(n), hw(n), c(n), g(n)}, realizou-se o seguinte
procedimento. Primeiramente, estimaram-se os cumulantes de 2ª, 3ª e 4ª ordem, baseado
respectivamente nas equações (4.71), (4.72) e (4.73). Assim, os índices de descorrelação
W2, W3,W4 foram calculados a partir de (4.74)-(4.75). Neste último caso, utilizou-se
τ
como um múltiplo inteiro de tempo de amostragem. A quantidade q, em (4.75), foi
ajustada a 10 vezes o tempo de amostragem, que é equivalente a aproximadamente 40
ms no caso de dados MEA, ou 1 ms no caso dos neurônios dos invertebrados.
A Tabela 5.1 apresenta a classificação dos sinais, baseado na previsão dos
resultados teóricos da literatura ((NIKIAS, 1993), (BONDON, 1991)), bem como em
experiência experimental prévia ((DESTRO-FILHO, 2001), (DESTRO-FILHO, 2002)
(DESTRO-FILHO, 2003)). Deve-se observar que, para valores dos índices de
descorrelação entre 0.30-0.74, a série temporal não é certamente nem Gaussiana, nem
periódica, nem pode ser classificada como descorrelacionada.
Tabela 5.1 - Classificação dos sinais baseados nos índices de descorrelação
Sinais W2,W3,W4
Ruído branco puro de ordem elevada
(não-gaussiano)
0.75-1.00
Correlacionado (não-gaussiano) 0.30-0.74
Periódico ou cíclico (não-gaussiano) 0.01-0.05
Gaussiano W3 > 23-25
W4 > 23-25

142
5.4 Resultados 1: EEG e Predição Linear
5.4.1. Processamento do banco de dados contendo sinais EEG com
crise
De forma a facilitar a análise dos resultados, foram construídos os seguintes
gráficos (Figuras 5.7 – 5.10), contendo os valores médios dos critérios utilizados (PM,
EQMr, TC e TE) em função da ordem de filtro (N).
Figura 5.7 - Gráfico PM x N para o processamento do banco de dados EEG/Crise.
Figura 5.8 - Gráfico EQMr x N para o processamento do banco de dados EEG/Crise.

143
Figura 5.9 - Gráfico TC x N para o processamento do banco de dados EEG/Crise.
Figura 5.10 - Gráfico TE x N para o processamento do banco de dados EEG/Crise.
Podem-se extrair as seguintes conclusões a partir dos gráficos acima:
PM: Apresenta como melhores ordens de filtro os valores compreendidos
entre 2 e 5. Nesse intervalo, os melhores passos de adaptação são 10
-6
e
10
-6
*exp(-n). Para os demais valores de N, o melhor passo de adaptação é
10
-6
*exp(-n);
EQMr: Apresenta menores valores para as ordens compreendidas entre 2 e 5.
Nesse intervalo, pode-se estabelecer os melhores passos de adaptação 10
-6
e 10
-5
;

144
TC: Os melhores valores de passo de adaptação foram 10
-6
*exp(-n) e 10
-5
. O
tempo médio de convergência é de 956,48 iterações;
TE: Os melhores valores de passo de adaptação foram 10
-6
e 10
-5
. O tempo
médio de estacionariedade é de 1289,32 iterações.
Observação: Os valores de N não foram descritos para os parâmetros TC e TE, visto
que estes se mantêm aproximadamente constantes. Portanto, pelo fato dos parâmetros
anteriores (
PM e EQMr) limitarem a análise ao intervalo de N de 2 a 5, assumem-se
estes valores como sendo os mais interessantes.
Conseqüentemente, um bom modelo paramétrico para o conjunto de sinais EEG
com crise pode ser obtido através de um filtro de predição de ordem N de 2 a 5,
utilizando-se passos de adaptação do tipo 10
-6
.
5.4.2. Processamento do banco de dados contendo sinais EEG sem
Crise
Os resultados desse processamento serão descritos pelos gráficos das Figuras
5.11 – 5.13, contendo os valores médios dos critérios utilizados (PM, EQMr e TC) em
função da ordem de filtro (N). Não se apresenta os gráficos de TE, pois não foi
encontrada nenhuma não-estacionariedade neste sinal.
Figura 5.11 - Gráfico PM x N para o processamento do banco de dados EEG/Sem Crise.

145
Figura 5.12 - Gráfico EQMr x N para o processamento do banco de dados EEG/Sem Crise.
Figura 5.13 - Gráfico TC x N para o processamento do banco de dados EEG/Sem Crise.
Podem-se extrair as seguintes conclusões acerca dos gráficos apresentados
acima:
PM: Para uma ordem de filtro 2<N<10, pode-se considerar como bom o passo
de adaptação de 10
-6
*exp(-n). Já para uma ordem de filtro de 7<N<10, o passo
de adaptação de 10
-5
é ótimo;

146
EQMr: Para uma ordem de filtro 7<N<10, pode-se considerar como bom o
passo de adaptação de 10
-5
. Para uma ordem de filtro 2<N<7, pode-se considerar
o passo de adaptação de 10
-6
e 10
-6
*exp(-n) como ótimo;
TC: Para uma ordem de filtro 3<N<8, o passo de adaptação de 10
-6
apresenta os
melhores resultados. Já para uma ordem de filtro 9<N<10, escolhe-se o passo de
adaptação de 10
-6
*exp(-n). O tempo médio de convergência é de 1082,3
iterações.
Conseqüentemente, um bom modelo paramétrico para o conjunto de sinais EEG
sem crise particular aqui usado pode ser obtido através de um filtro de predição de
ordem N de 2 a 7, utilizando-se passos de adaptação do tipo 10
-6
.
5.4.3. Comparação dos resultados do banco de dados de EEG/Crise e
EEG/ Sem Crise
As Tabelas 5.2, 5.3 e 5.4 apresentam a comparação dos resultados obtidos nos
processamentos dos bancos de dados de sinal EEG/Crise e EEG/Sem Crise.
Tabela 5.2 - Comparação dos Resultados EEG/Crise e EEG/Sem Crise:
Ordem de filtro e passo de adaptação.
EEG/Crise EEG/Sem Crise
Ordem do Filtro
2 a 5 2 a 7
Passo de Adaptação
10
-6
10
-6
Tabela 5.3 - Comparação dos Resultados EEG/Crise e EEG/Sem Crise:
Tempo de convergência e tempo de estacionariedade.
EEG/Crise EEG/Sem Crise
Em iterações Em segundos Em iterações Em segundos
TC
956,48 4,78s 1082,3 5,41s
TE
1289,32 6,45s - -

147
Tabela 5.4 - Comparação dos Resultados EEG/Crise e EEG/Sem Crise:
Pico máximo e erro quadrado médio de regime permanente.
EEG/Crise EEG/Sem Crise
PM
3,3052*10
5
5,1301*10
6
EQMr
1,3332*10
4
1,5486*10
3
A Tabela 5.2 evidencia resultados muito semelhantes no que diz respeito aos
valores de filtro de predição e de passo de adaptação para os sinais EEG/Crise e
EEG/Sem Crise.
Os valores das Tabelas 5.3 e 5.4 foram obtidos através da média aritmética
tomada sobre a ordem 5, e considerando o passo de adaptação 10
-6
.
A Tabela 5.3 apresenta resultados distintos, embora próximos, para o TC em
ambos os casos. Porém, não foram especificados valores de TE para o sinal EEG/Sem
Crise, visto que não foram observadas não-estacionariedades.
Na Tabela 5.4, pode-se observar que os valores de PM e EQMr diferem quanto à
ordem de grandeza, se comparados entre diferentes sinais. Contudo, pode-se concluir
que a previsão de um sinal sem crise epiléptica é mais precisa.
Logo, um bom modelo paramétrico para o conjunto de sinais EEG com e sem
crise pode ser obtido através de um filtro de predição de ordem 2 a 5, utilizando-se
passos de adaptação do tipo 10
-6
.
5.5 Resultados 2: MEA e Predição Linear
5.5.1. Processamento de dados de sinais MEA – Teste 1
Os resultados desse processamento serão descritos pelos gráficos das Figuras
5.14 – 5.16, contendo os valores médios dos critérios utilizados (PM, EQMr e TC) em
função da ordem de filtro (N). Utilizou-se um único sinal MEA- Basale 1 (associado a
um único eletrodo), contendo 3.10
6
amostras, as quais foram subdivididas em 1000
arquivos de 3000 pontos cada um. Os resultados apresentados correspondem à média de
todos esses arquivos. Para este sinal MEA não foi encontrado nenhuma não-
estacionariedade, portanto não se discute o TE.

148
Figura 5.14 - Gráfico PM x N para o sinal de MEA.
Figura 5.15 - Gráfico EQMr x N para o sinal de MEA.

149
Figura 5.16 - Gráfico TC x N para sinal de MEA.
Podem-se extrair as seguintes conclusões a partir dos gráficos acima:
PM: Apresenta como melhores ordens de filtro os valores menores que 50, ou
seja, N < 50. Nesse intervalo, o melhor passo de adaptação é 10
-6
;
EQMr: Apresenta menores valores para as ordens menores que 20 (N<20).
Nesse intervalo, pode-se estabelecer os passos de adaptação 10
-6
e 10
-5
como
ótimos;
TC: Os melhores valores de passo de adaptação foram 10
-6
e 10
-5
e ordens
menores que 20. O tempo médio de convergência é de 2645,5 iterações;
Observação: Pode-se observar que, para o passo de adaptação 10
-4
, o filtro com ordens
superiores a 20 não converge, ou seja, ele nem não consegue realizar a predição. Para as
ordens maiores que 20, qualquer que seja o passo de adaptação, o
TC é muito elevado.
Portanto, um bom modelo paramétrico para o conjunto de sinais MEA analisado
pode ser obtido através de um filtro de predição de ordem N ≤ 20, utilizando-se passos
de adaptação do tipo 10
-6
. Consequentemente, deve-se refinar a análise para N<20 e
passos de adaptação da ordem 10
-6
e 10
-5
, para melhores resultados na análise do filtro
de predição. Isso é mostrado na seção a seguir.

150
5.5.2. Processamento de dados de sinais MEA – Teste 2
Os resultados desse processamento serão descritos pelos gráficos das Figuras
5.17 – 5.19, contendo os valores médios dos critérios utilizados (PM, EQMr e TC) em
função da ordem de filtro (N).
Figura 5.17 - Gráfico PM x N para o sinal de MEA.
Figura 5.18 - Gráfico EQMr x N para o sinal de MEA.

151
Figura 5.19 - Gráfico TC x N para sinal de MEA.
Podem-se extrair as seguintes conclusões a partir dos gráficos acima:
PM: Apresenta como melhores ordens de filtro os valores menores que 5, ou
seja, N < 5. Nesse intervalo, os melhores passos de adaptação são 10
-6
e 10
-5
;
EQMr: Apresenta menores valores para as ordens menores que 5 (N<5). Nesse
intervalo, pode-se estabelecer os passos de adaptação 10
-6
e 10
-5
, como ótimos;
TC: O melhor valor de passo de adaptação foi 10
-6
, para ordens de filtro
menores que 4 (N<4). O tempo médio de convergência é de 2477,8 iterações;
Observação: Os comportamentos do filtro com os passo de adaptação 10
-6
*exp(-n) e
10
-6
/n são muito semelhantes.
Consequentemente, um bom modelo paramétrico para o conjunto de sinais MEA
analisados pode ser obtido através de um filtro de predição de ordem N <5, utilizando-
se passos de adaptação do tipo 10
-6
e 10
-5
.
5.5.3. Resumo dos resultados dos Testes 1 e 2 e comparação dos
resultados obtidos com os sinais EEG
As Tabelas 5.5, 5.6 e 5.7 apresentam a comparação dos resultados obtidos nos
processamentos do sinal MEA, onde os valores das Tabelas 5.6 e 5.7 são obtidos através

152
da média aritmética tomada sobre todas as ordens menores que 10, e considerando o
passo de adaptação 10
-6
.
Tabela 5.5 - Comparação dos Resultados da MEA:
Ordem de filtro e passo de adaptação.
MEA
Ordem do Filtro
N<5
Passo de Adaptação
10
-6
Tabela 5.6 - Comparação dos Resultados MEA:
Pico máximo e erro quadrado médio de regime permanente.
MEA
PM
1,95204*10
5
EQMr
84,82
Tabela 5.7 - Comparação dos Resultados MEA:
Tempo de convergência.
MEA
Em iterações Em segundos
TC
2477,8 9,91s
Analisando-se os resultados finais da predição, resumidos pelas Tabelas 5.2-5.4
e 5.5-5.7, concluímos que este modelo funciona adequadamente tanto para os sinais
MEA (nível celular) como para sinais EEG (nível neurológico). Além disso, os valores
das ordens de filtro e passo de adaptação são muito próximos.
Observa-se também os valores de PM são próximos, todavia o modelo de sinal
MEA é mais estável, pois apresenta um pico menor que aquele do sinal EEG. Por outro
lado, o modelo de MEA também está associado a valores menores do EQMr e, portanto,
é mais preciso que para os outros dois sinais EEG.

153
Com relação ao TC, conclui-se que, para os sinais EEG, os resultados são muito
próximos. Já para o sinal MEA, TC é muito elevado, se comparado com os sinais EEG,
tornando o modelo muito lento para processamento em tempo real.
5.6 Resultados 3: MEA como Ruído Branco
As Tabelas 5.8 - 5.11 descrevem os resultados que foram obtidos após o cálculo
dos índices IDE, realizado a partir da média realizada sobre 100 seqüências de cada
sinal, onde cada seqüência analisada tem um comprimento de 40.000 amostras. Na
Tabela 5.8, pw(n) representa a definição teórica do ruído branco puro de ordem elevada,
como estabelecido em (4.69)-(4.70).
Tabela 5.8 – Índices de descorrelação para diversos sinais
Sinais
W2 W3 W4
i(n)
0.85 0.85 0.82
hw(n)
0.90 0.90 0.85
pw(n)
1.00 1.00 1.00
c(n)
0.04 0.03 0.02
g(n)
25 24 23
Na Tabela 5.8, os índices de descorrelação associados com as definições hw(n),
c(n) e g(n) representam, respectivamente, os limites práticos da amplitude de IDE que
caracterizam a série temporal descorrelacionada, a atividade periódica e os processos
estocásticos gaussianos. Estes limites são de grande importância, pois permitem a
classificação dos sinais como ruído branco, cíclico ou (não) gaussiano. Deve-se
observar na Tabela 5.8 que os índices de descorrelação experimental para os sinais
hw(n), c(n) e g(n) concordam com os resultados apresentados na Tabela 5.1.
A Tabela 5.8 indica que a atividade espontânea dos sinais de invertebrados é
certamente um sinal descorrelacionado, visto que os índices de descorrelação são muito
próximos aos índices associados com o ruído branco de ordem elevada hw(n).
As Tabelas 5.9, 5.10 e 5.11 descrevem a classificação dos sinais registrados em
diversos canais da MEA. É interessante observar que os resultados permitem a

154
classificação dos sinais de acordo com ambas as Tabelas 5.1 e 5.8. Além disso, a
maioria dos sinais MEA são correlacionados. Quatro canais apresentam um
comportamento do tipo ruído branco (Tabela 5.9) e apenas dezoito canais podem ser
classificados como atividade cíclica (Tabela 5.11); enquanto que 38 canais são
classificados como correlacionados (Tabela 5.10). Deve-se destacar que os sinais
periódicos ocorrem em locais de eletrodos vizinhos da MEA.
Tabela 5.9 – IDE para os sinais MEA descorrelacionados
Número dos
Canais MEA
W2 W3 W4
1, 58 0.82 0.80 0.79
10, 46 0.87 0.81 0.80
Tabela 5.10 – IDE para os sinais MEA correlacionados
Número dos Canais
MEA
W2 W3 W4
2, 34 0.53 0.50 0.47
7, 38 0.53 0.52 0.51
8, 30 0.50 0.47 0.45
12, 55 0.52 0.51 0.50
13, 45 0.55 0.54 0.52
14, 20 0.49 0.49 0.47
15, 31 0.62 0.61 0.59
21, 59 0.63 0.63 0.59
27, 33 0.53 0.53 0.52
42, 54 0.51 0.50 0.47
47, 23 0.50 0.49 0.47
11, 56 0.55 0.54 0.53
16, 32 0.67 0.66 0.65
22, 53 0.50 0.49 0.48
24, 57 0.57 0.56 0.55
25, 39 0.59 0.59 0.58
60, 28 0.53 0.52 0.51
37, 29 0.54 0.53 0.52
41, 52 0.60 0.59 0.58

155
Tabela 5.11 – IDE para os sinais MEA cíclicos
Número dos
Canais MEA
W2 W3 W4
3, 4, 5, 6 0.09 0.07 0.05
9, 17, 18, 19 0.10 0.07 0.05
26, 35 0.08 0.04 0.02
36, 40 0.09 0.09 0.08
43, 44, 48, 49 0.08 0.06 0.05
50, 51 0.07 0.06 0.04
Finalmente, a observação de todas as Tabelas 5.8-5.11 permite concluir que
todos os índices de descorrelação apresentam valores limitados, ou, ao menos, valores
pequenos se comparados com os índices IDE associados à série temporal Gaussiana
g(n) (veja a Tabela 5.8). Consequentemente, todos os sinais de atividade espontânea
podem ser considerados processos estocásticos não-gaussianos.
5.7 Conclusão
Em resumo, pode-se ressaltar as seguintes conclusões para sinais EEG:
1) O filtro de predição que modela o banco de dados EEG deve ter no máximo
comprimento
, para o caso do sinal com crise ou sem crise. Os passos de
adaptação devem estar ajustados para valores próximos a 10
5=N
-6
, incluindo-se a
possibilidade de passo variável do tipo exponencial.
2) É possível realizar predição linear de excelente qualidade, em particular de trechos de
sinal com crise. O ajuste do erro quadrático médio permite melhorar o desempenho do
preditor no caso de sinais com crise, o que irá refletir em um menor valor para o pico
máximo.
3) Analisando os resultados do sinal EEG (crise e sem crise), percebe-se que os valores
de ordem de filtro e passo de adaptação são muito semelhantes. Já os valores de TC são
distintos, mas próximos. O sinal sem crise não apresenta não-estacionariedades. Já para
os valores de PM e EQMr, estes são diferentes quanto à ordem de grandeza. A previsão
de um sinal sem crise é mais precisa e estável do que aquela de um sinal com crise
epiléptica.
156
4) Observando-se os sinais EEG com e sem crise epiléptica, pode-se comparar os
tempos médios de convergência e de estacionariedade, o que permite concluir o
seguinte. Um neuroimplante que se baseia na técnica de predição linear para a previsão
de crises epilépticas não poderá realizar uma previsão minimamente razoável antes de
5,41 segundos, e também não poderá antecipar a previsão da crise além de 6,45
segundos. Deve-se observar que não foram constatados saltos não-estacionários para os
trechos de sinal sem crise.
5) Durante a ocorrência de uma crise epiléptica do lobotemporal, alterações estatísticas
abruptas acontecem no sinal de EEG, em média, a cada 6,45 segundos. Este tempo de
estacionariedade poderia estar associado a algum fenômeno fisiológico, responsável por
alterações neurodinâmicas.
Do ponto de vista de sinais MEA, pode-se concluir que:
1) O filtro de predição que modela o sinal da MEA deve ter no máximo comprimento
. O passo de adaptação deve estar ajustado para valores próximos de 105=N
-6
.
2) Pode-se observar que para valores com ordens de filtro muito grandes, ou seja, N
>50, ele não converge para o passo de adaptação 10
-4
. Desta maneira, para estas mesmas
ordens, qualquer que seja o passo de adaptação, o valor de TC é muito alto, tornando o
filtro muito lento para convergir. É importante ressaltar que os passos de adaptação
10
-6
*exp(-n) e 10
-6
/n conduzem a um desempenho semelhante.
3) O modelo do sinal MEA exige aproximadamente 10 segundos para ser estimado.
Todavia, não se observou nenhuma não-estacionariedade deste sinal.
4) Analisando o modelo auto-regressivo dos sinais EEG (crise e em crise), conclui-se
que ele converge rapidamente, é instável e grosseiro, se comparado com o modelo do
sinal MEA; já que este último converge lentamente, ao mesmo tempo que é estável e
bem mais preciso. Portanto, o sinal EEG possui uma variação estatística maior que
aquela do sinal MEA, daí a dificuldade em se estimar a predição em tempo real.
5) Estabeleceram-se limites experimentais para os índices de descorrelação IDE de 2ª,
3ª e 4ª ordem, de modo a classificar os sinais de atividade neural espontânea, como
ruído branco de ordem elevada, gaussiano de ordem elevada, ou (des) correlacionado de
ordem elevada. Estes limites concordam com os resultados precedentes na literatura
157
((NIKIAS, 1993), (DESTRO-FILHO, 2002), (DESTRO-FILHO, 2003), (BONDON,
1991)).
6) Particularmente, visto que a atividade espontânea dos neurônios dos invertebrados é
certamente um ruído de ordem elevada, os resultados confirmam as conclusões de
(BLESIC, 1999).
7) Os sinais MEA são, em grande maioria, correlacionados; ocorrendo uma atividade
periódica em eletrodos vizinhos. Todos os sinais de atividade espontânea analisados são
não-gaussianos, satisfazendo, portanto a condição principal para aplicação das
estatísticas de ordem elevada como técnica de processamento.
Finalmente, deve-se ressaltar que as conclusões acima resumidas são restritas
aos bancos de dados analisados.
158
Capítulo 6
Conclusões e Trabalhos Futuros
6.1 Conclusões gerais
Este capítulo retoma algumas das questões dos modelos simples do ponto de
vista dos resultados obtidos ao longo do trabalho, da adequação aos desafios iniciais da
dissertação, e do interesse para investigações futuras.
As novas terapias referem-se aos desafios que constam nesta dissertação. Deve-
se destacar o desenvolvimento de ferramentas teóricas ao contexto não-estacionário dos
sinais e à não-lineariedade dos sistemas. Com isso, desenvolveram-se algoritmos de
baixa complexidade, permitindo operação em tempo real, visto que o sistema deve
processar, simultaneamente, através de MEAs, tanto sinais neurobiológicos (biológicos
em nível celular) quanto em nível de massa neural encefálica (neurológico). Em longo
prazo, o estabelecimento de conexões entre as informações ligadas aos dois níveis
biológicos permitirá uma melhor compreensão do funcionamento do sistema nervoso
humano.
Uma vez recordados os conceitos fundamentais em neurofisiologia e neurologia,
foram introduzidas novas técnicas de medidas neurofisiológicas, em particular as
matrizes multieletrodo (MEA) para aplicações em neuroimplantes. Esta tecnologia, que
corresponde a uma evolução do conceito de interface bioeletrônica, é extremamente
159
cara e ainda está longe de atingir o grau de confiabilidade e biocompatibilidade exigidos
para aplicações clínico-terapêuticas. Todavia, ela se presta atualmente a estudos
fundamentais para a compreensão da neurodinâmica. De qualquer forma, o
desenvolvimento da MEA, aliada ao rompimento de barreiras no modelamento de
sistemas complexos, tende em longo prazo estabelecer novos pontos de vista sobre
patologias do sistema nervoso.
Dentre essas patologias, destaca-se a epilepsia, um problema considerado grave,
além de representar um desafio da saúde pública mundial, em termos de controle e
prevenção. Atualmente, 60 milhões de pessoas são portadores da doença, destacando-se
os pacientes que são tratados por drogas anti-epilépticas, outros cirurgicamente, e ainda
os casos refratários a qualquer tratamento existente.
Em particular, pode-se também utilizar matrizes multieletrodo em projetos de
microssistemas para testes de drogas, e neste caso as respostas de células a estímulos
químicos são avaliadas em função da droga aplicada. Para aplicações em implantes
neurais ou neuromusculares crônicos, são usualmente propostas estruturas em
multicamadas, que viabilizam o estímulo e a aquisição de sinais extracelulares, como
por exemplo, do nervo óptico.
De acordo com a literatura, as ferramentas do processamento de sinais
atualmente aplicadas para a análise do EEG apresentam elevada complexidade
computacional, que é contraditório com as exigências de biocompatibilidade impostas
pela implementação de neuroimplantes. Já para o caso de sinais MEA, as técnicas em
uso dificultam a operação em tempo real.
Dentre as técnicas analisadas para superar as limitações da literatura, abordadas
no parágrafo anterior, destacam-se a predição linear adaptativa e a estatística de ordem
elevada (EOE), que constituem ferramentas adequadas ao contexto de sinais não-
estacionários e sistemas não-lineares.
O uso de filtros adaptativos permite eliminar as interferências durante a
aquisição dos sinais, particularmente para processamento em tempo real, onde não se
conhece a estatística do sinal a ser analisado. Dentre as diversas técnicas de
processamento digital de sinais (PDS), destaca-se a predição linear, que permite um
bom desempenho através de algoritmos simples e de baixa complexidade
160
computacional. Por outro lado, a teoria de EOE é muito importante para os estudos em
engenharia neural, fornecendo informações precisas sobre a função de densidade de
probabilidade, permitindo uma análise mais exata do sinal. Contudo, não se pode
esquecer das limitações da teoria de EOE, destacando-se o alto custo computacional e a
lentidão de convergência de seus estimadores. Muito embora tais limitações possam ser
consideradas contrárias aos desafios do Capítulo 2, o uso de técnicas EOE possibilita
usufruir de propriedades ligadas aos cumulantes, como por exemplo, a imunidade ao
ruído gaussiano, sempre presente em medidas eletrofisiológicas, e a especificidade de
caracterização de sistemas não-lineares. Em particular, neste último intuito, a
dissertação enfocou o uso dos índices de descorrelação elevada.
Os resultados obtidos através de simulações desta dissertação permitem concluir
que:
O filtro de predição linear funciona adequadamente para ambos os sinais
processados, sendo que para o sinal MEA, ele é mais estável e preciso. Em
termos dos sinais EEG, a predição linear de trechos sem crise é mais precisa e
estável. Para qualquer tipo de sinal considerado nesta dissertação, a ordem do
filtro deve ser menor que N=5, e o passo de adaptação deve assumir valores
entre 10
-5
e 10
-6
.
Os valores dos parâmetros do filtro são distintos, mais próximos. Uma
observação importante é quanto ao tempo de convergência, que para sinais de
EEG foi bem mais rápido que o sinal de MEA.
Contudo, não se observou nenhuma não-estacionariedade nos sinais de MEA e
EEG sem crise epiléptica, enquanto este fenômeno foi constatado muito
claramente no sinal de EEG com crise. Consequentemente, o grau de variação
da estrutura estatística dos sinais EEG é mais intenso que o grau do sinal MEA,
relativamente à variável tempo.
Analisando-se os sinais EEG com e sem crise, conclui-se que um neuroimplante
baseado em predição linear não poderá realizar uma previsão minimamente
razoável antes de 5,41 segundos, e não antecipará uma crise além de 6,45
segundos. Além disso, não é possível se fazer uma previsão minimamente
razoável de um sinal MEA antes de 10 segundos.
161
Em conseqüência da conclusão anterior, e fundamentado pelo conceito de
“tempo de estacionariedade”, pode-se afirmar que o janelamento ótimo de
sinais EEG corresponde a aproximadamente trechos de 6 segundos, enquanto
que, para sinais MEA, estes trechos correspondem a 10 segundos. Além disso,
pode-se também concluir que alterações neurodinâmicas acontecem, em média,
a cada 6 segundos durante a ocorrência de crises epilépticas do lobotemporal.
Os limites experimentais dos índices de descorrelação IDE de 2ª, 3ª e 4ª ordem,
para classificação de sinais de atividade neural espontânea, concordam com os
resultados da literatura.
Foi observado que a maior parte dos sinais de MEA são correlacionados,
ocorrendo uma atividade periódica em eletrodos vizinhos. Para sinais de
atividade espontânea associados a um pequeno grupo de neurônios, as séries
temporais são claramente do tipo “ruído branco”, em conformidade com a
literatura.
A condição principal de uso da teoria de EOE, como técnica de processamento,
foi verificada para todos os sinais da MEA, que podem ser considerados como
não-gaussianos. Consequentemente, a utilização deste ferramental abre novas
perspectivas para o estudo da codificação neural.
Em resumo, os resultados demonstram que é possível realizar uma boa predição
de um sinal biológico não-estacionário, seja num contexto onde se exige tempo real
(EEG), seja em outro contexto (sinal MEA). Além disso, as técnicas baseadas na teoria
de EOE podem ser usadas com sucesso para sinais MEA, para os quais, pelo menos no
contexto atual das culturas in vitro, não se exige processamento em tempo real. Isto
justifica a aplicação da teoria de EOE, caracterizada como lenta e complexa. Estes
modelos, baseados na predição linear e no ruído branco, tornam-se importantes
ferramentas para a Medicina, no que diz respeito às novas terapias, em particular a
prevenção de crises epilépticas. Desta forma, pode-se cogitar o desenvolvimento de
neuroimplantes, baseados em sinais MEA. Estes modelos também representam
importante contribuição para a Biologia, como instrumentos de análise para refinar a
compreensão de fenômenos fisiológicos, tanto em nível celular, como em nível do
córtex cerebral.
162
6.2 Trabalhos futuros
Como possível seqüência deste trabalho, pode-se citar:
Aprofundamento da pesquisa bibliográfica associada ao Capítulo 3;
Realização da Predição Linear Adaptativa incluindo todo o banco de dados de
MEA, e não apenas para alguns eletrodos, além da possível utilização do passo
de adaptação variável;
Aplicação do Método de Segmentação Adaptativa (COHEN, 2000) para os
sinais EEG e MEA, com o objetivo de se calcular o tempo de estacionariedade e
dimensionar um janelamento ótimo;
Estimativa das correlações de ordem elevada até a 7ª ordem estatística, para
validar os resultados atuais e aqueles descritos em ((DESTRO-FILHO, 2002),
(DESTRO-FILHO, 2003)). Em particular, as correlações de ordem elevada
devem ser recalculadas após a segmentação dos sinais de atividade espontânea,
para refinar a estimação dos índices de correlação realizada por esta dissertação;
Aprofundamento do significado fisiológico sobre do conceito de tempo de
estacionariedade, em particular, sobre sua associação com achados clínicos
ligados à epilepsia.
163
Referências Bibliográficas
1 AERTSEN, A. M. H. J.; GERSTEIN, G. Evaluation of neuronal connectivity:
sensitivity of cross-correlation,
Brain research, 340, p. 341-354, 1985.
2 AKAY, M.
Nonlinear biomedical signal processing. Prentice-Hall Inc, USA,
356 p., 2003.
3 BANKER, G.; GOSLIN, K.
Culturing Nerve Cells. MIT Press, 1991.
4 BARDAKJIAN, B.L.; et al. Nonlinear system identification of hippocampal
neurons. In: MARMARELIS, V.Z. (ed.).
Advanced Methods of Physiological
Systems Modeling.
New York: Plenum Press, v.3, p. 179-194, 1994, 2003.
5 BAY, Q.; WISE, K.D.; ANDERSON, D.J. A high-yield microassembly
structure for three-dimensional microelectrode arrays.
IEEE Transactions on
Biomedical Engineering
, v.47, n.3, p. 281-289, 2000.
6 BEADLE, D.J.; LEES, G.; KATER, S.B.
Cell culture approaches to
invertebrate neuroscience.
London: Academic Press, 1988.
7 BERGER, T. W.; et al. Brain-Implantable biomimetic electronics as the next
era in neural prosthetics.
Proceedings of the IEEE, Special Issue on
Neuroengineering
, v.89, n. 7, 2001.
8 BERNARDI, A.
Filtros de Kalman no Levantamento de Características do
Sinal de EEG
. 1999. 157f. Dissertação (Mestrado). Escola Federal de
Engenharia de Itajubá, Itajubá, 1999.
9 BLANCO, S.; et al. On the stationary of EEG signals.
IEEE Engin. In
Medicine and Biology Magazine
, p. 395-399, July-August 1995.
10 BLESIC, S.; MILOSEVIC, S.; et al. Detrended fluctuation analysis of time
series of a firing fusimotor neuron.
Physica A, v. 268, p. 275-282, 1999.

164
11 BLESIC, S.; MILOSEVIC, S.; STRATIMIROVIC, Dj.; LJUBISAVLJEVIC,
M. Detecting long-range correlations in times series of neuronal discharges.
Physica A, v. 330, n. 3, p. 391-399, Dec. 2003.
12 BOHTE, S.M.; et al. the effects of pair-wise and higher-order correlations on
the firing rate of a postsynaptic neuron.
Neural Computing, v. 12, p. 153-179,
2000.
13 BONDON, P.; PICINBONO, B. Whiteness and its transformations.
Traitement
du Signal
, v.7, n. 5, p. 385-395, Dec. 1991.
14 BONIFAZI, P.; FROMHERZ, P. Silicon chip for electronic communication
between nerve cells by non-invasive interface and analog-digital processing.
Advanced Materials, v.14, n.17, p. 1190-1193, Sept. 2002.
15 BORST, Alexander; THEUNISSEN, Frederic E. Information theory and neural
coding.
Nature Neuroscience, v. 2, nº 11, p. 947-957, Nov. 1999.
16 BOVE, M.; GRATTAROLA, M.; VERRISCHI, G. In vitro 2-D networks of
neurons characterized by processing the signals recorded with a planar
microtransducer array.
IEEE Transactions on Biomedical Engineering, v.40,
n.10, p. 964-977, 1997.
17 BRAY, D. Surface movements during the growth of single explanted neurons.
Proc. Nat. Acad. Sci. USA, v. 65, p. 905-910, 1970.
18 BRINDLEY, GS; LEWIN, WW. The sensations produced by electrical
stimulation of the visual cortex.
J. Physiol., v. 196, p. 479-493, 1968.
19 BUTTON, V. L. S. N. Eletroencefalógrafos e monitores de EEG. In
: IA748
Instrumentação Biomédica
. Campinas: DEB/FEEC – Unicamp, 2000.
Disponível em: <
www.deb.fee.unicamp/eeg.pdf>. Acessado em: 28 ago. 2005.
20 CADZOW, J. A. Blind Deconvolution via Cumulant Extrema.
IEEE Signal
Processing Magazine
, p. 24-42, May 1996.
21 CARDOSO, R. R.
Uma estratégia de modelagem tridimensional para
mapeamento de EEG de superfície
. 2005. 159f. Dissertação (Mestrado).
Universidade Federal de Uberlândia, Uberlândia, 2005.
22 CARDOSO, R. R.
Filtragem de Kalman aplicada ao processamento de
sinais eletroencefalográficos (EEG)
. Relatório técnico, FEELT/UFU, Julho,
2005.
23 CASTELLONE, P.; RUTTEN, W.L.C.; MARANI, E. Cross-interval histogram
analysis of neuronal activity on multi-electrode array.
Proceedings of the 1
st
165
International IEEE EMBS Conference on Neural Engineering, Capri Island,
Italy, p. 301-304, March 2003.
24 CHAN, Chiu-Hsien; SRINIVASAN, Vijay; HSIAO, Min-Chi; KHANNA,
Sharad; WILLS, Jack D.; GHOLMIEH, Ghassan; LaCROSS, Jeff;
COURELLIS, Spiros; Granacki, John J.; BERGER, Theodore. Real Time
Hardware Neural Spike Amplitude Extraction.
IEEE International Workshop
on Biomedical Circuits and Systems
, S.l.8, p. 20-23, 2004.
25 CHAPIN, JK; MOXON, KA; MARKOWITZ, RS; NICOLELIS, MA. Real-
time control of a robot arm using simultaneously recorded neurons in the motor
cortex.
Nat. Neuroscience, v.2, n.7, p. 664-670, July 1999.
26 CHIAPPALONE, M.; et al. Networks of neurons coupled to microelectrode
arrays: a neuronal sensory system for pharmacological applications.
Biosensors
and Bioelectronics
, v.18, p. 627-634, 2003.
27 COCKERELL, O.C.; SANDER, J.W.A.S. O custo Econômico da Epilepsia. In:
COSTA, J.C. da et al. (ed).
Fundamentos Neurobiológicos das Epilepsias.
São Paulo: Lemos-Editorial, p. 21-30, 1998.
28 COHEN, A.
Biomedical Signal Processing. v.1: Time and Frequency
Analysis; v.2: Compression and Classification, CRC Press, Boca Raton, FI,
USA, 1986.
29 COHEN, A.
Biomedical Signal Processing. Volume I. Time and Frequency
Domains Analysis.
Florida: CRC Press, 2000.
30 COHEN, A.
Biomedical Signal Processing. Volume II. Compression and
Automatic Recognition.
Florida: CRC Press, 2000.
31 COZZI, L.; ANGELO, P.D; CHIAPPALONE, M.; IDE, A. N.; NOVELLINO,
A.; MARTINOIA, S.; SANGUINETI, V. Coding and decoding of information
in a bi-directional neural interface.
Elsevier Science Neurocomputing, v. 65-
66, p.783-792, 2005.
32 D’ALESSANDRO, M.; et al. A multi-feature and multi-channel univariate
selection process for seizure prediction.
Clinical Neurophysiology, v. 116, p.
506-516, 2005.
33 DE FATTA, D.J.; LUCAS, J.G.; HODGKISS, W.S.
Digital Signal Processing:
a system design approach
. New York: Wiley, 1988.
34 DESTRO-FILHO, J.B.; PEIXOTO, N.; et al. On the relationships between
Hebbian synaptic plasticity, (Non)linear prediction and High-Order Statistics.
Proc. Of IPCAT’2001, p. 43-54, Leuvain, Bélgica, 13-17 August 2001.
35 DESTRO-FILHO, J.B.; PEIXOTO, N.; BLESIC, S. High-order statistical
166
analysis of neuronal coding: case study on the “whiteness” of spontaneous
activity.
Proc. of IEEE/EURASIP BIOSIGNAL’2002, p. 162-164, Brno,
República Checa, May 2002.
36 DESTRO-FILHO, J.B. High-order whiteness and its application to
telecommunications.
Proc. of IEEE Workshop NSIP’03, Grado, Itália, 08-11,
June 2003.
37 DURAND, D. M.; BIKSON, M. Supression and control of Epileptiform
Activity by Electrical Stimulation: A Review.
Proc. of the IEEE, vol.89, n.7,
p.1065-1082, July 2001.
38 EGERT, U.; NISCH, W.; HAMMERLE, H. Simultaneous recording of the
electroretinogram and spike activity with a microelectrode-array.
18
th
Annual
International Conference of the IEEE Engineering in Medicine and
Biology Society
, Amsterdam, 1996.
39 EICHLER, Michael; DAHLHAUS, Rainer; SANDKUHLER. Jurgen. Partial
correlation analysis for the identification of synaptic connections.
Biological
Cybernetics
, v. 89, p. 289-302, Out. 2003.
40 FRISÓN, J.; et al.
Enciclopédia do Corpo Humano. São Paulo: Editorial
Planeta-DEAgostini S.A., 1999.
41 FROMHERZ, P. Self-focusing on ion channels in cell adhesion.
Physical
Review E
, v.52, n.2, p. R.1303-R1305, 1994.
42 FROMHERZ, P.; STETT, A. Silicon-Neuron junction capacitive stimulation of
an individual neuron on a silicon chip.
Phys. Rev. Lett., v. 75, p. 1670-1673,
1995.
43 FROMHERZ, P.; BRAUN, D. Fluorescence interference-contrast microscopy
of cell adhesion on oxidized silicon.
Applied Physics A: Materials Science &
Processing
, v. 65, n.4-5, p. 341-348, 1997.
44 FROMHERZ, P. Neuroelectronic Interfacing: Semiconductor Chips with Ion
channels, Nerve Cells, and Brain.
In: Nanoelectronics and Information
Technology
, Berlin, p. 781-810, 2003.
45 GERSTEIN, G. L.; MANDELBROT, B. random walk models for the spike
activity of a single neuron,
Biophys. J., v.4, p. 41-68, 1964.
46 GIMENEZ, J.R.B.
Sobre a estabilidade numérica dos algoritmos de
mínimos quadrados rápidos
. 1995. 160f. Tese (Doutorado), Universidade
Estadual de Campinas, São Paulo, 1995.
47 GLASER, E.M.; RUCHKIN, D.S.
Principles of neurobiological signals
analysis.
Academic Press, New York: USA, 471pp., 1976.
167
48 GRILL, WM; KIRSCH, RF.
Neural prostheses. In: Wiley Encyclopedia of
Electrical and Electronics Engineering, ed. JG Webster, p. 339-350. New York:
Wiley, 1999.
49 GROSS, G.W.; RHOADES, B.K.; REUST, D.L.; SCHWALM, F.U.
Stimulation of monolayer networks in culture through thin-film indium-tin
oxide recording electrodes.
J. Neuroscience Methods, v.50, p. 131-143, 1993.
50 GUEDES, L.A., BAKER, L.E.
Principles of applied biomedical
instrumentation
, New York: Wiley, 1989.
51 GUYTON, A. C. Tratado de Fisiologia Médica. 10ª ed. Rio de Janeiro:
Editora Guanabara Koogan S.A., 2002.
52 HAIDARLIU, S.; SHULZ, D.; AHISSAR, E. A multi-electrode array for
combined microiontophoresis and multiple single-unit records.
J. Neuroscience
Methods
, v.56, p. 125-131, 1995.
53 HAYKIN, S.
Adaptive Filter Theory, 2nd Edition. USA: Prentice-Hall, 1991.
54 HAYKIN, S., (ed.).
Unsurpervised adaptive filtering. USA: Jonh Wiley and
Sons Inc, vol. 1-2, 2000.
55 HILTUNEN, J.K.; KAIPIO, J.P. Comparison of the accuracy of two
segmentation methods in medium rate EEG transitions.
Proceedings of the
22
nd
Annual EMBS International Conference, p. 3018-3021, July 2000.
56 HOLLAND, N. D. Early Central Nervous System evolution: an era of skin
brain?
Nature Reviews Neuroscience, v.4, p. 617-627, Aug. 2003.
57 HUAI-YUAN, C.; TZU-YING, K.; BAOWEN, C.; SHAO-WEI, L.; CHUAN-
CHIN, C.; WEILEUN, F. Development of the three dimensional multi-
electrode array for neural recording.
The 13
th
International conference on
Solid-State Sensors, Actuators and Microsystems
, Korea, June 2005.
58 IASEMIDIS, L.D. Epileptic seizure prediction and control.
IEEE Transactions
on Biomedical Engineering
, Special Issue on Epileptic Seizure Prediction,
vol.50, n.5, p. 549-563, May 2003.
59 ISRAEL, D.A.; BARRY, W.H.; EDELL, D.J.; MARK, R.G. An array of
microelectrodes to stimulate and record from cardiac cells in culture.
American
Journal of Physiology
, v.247, p.H669-H674, 1984.
60 IZHIKEVICH E. M. Simple Model of Spiking Neurons. IEEE Transactions on
Neural Networks, v.14, n. 6, p. 1569-1572, Nov. 2003.
61 KANDEL, E. R.; SCHWARTZ, J. H.; JESSEL, T. M.
Fundamentos da

168
Neurociência e do Comportamento. Rio de Janeiro: Editora Guanabara
Koogan AS, 2000.
62 KRAUSE, M.; WHITSON, J.; et al.
Accelerating Electrophysiology using
higher-throughput multielectrode array.
Recent Advances in Network
electrophysiology using multi-electrode arrays.
Convention Center, New
Orleans, LA, Nov. 2003.
63 LATHI, B.P.
An Introduction to Random Signals. USA: John Wiley and
Sons Inc., 1967.
64 LENT, R.
Cem Bilhões de Neurônios: conceitos fundamentais de
neurociência
. São Paulo: Editora Atheneu, 2001.
65 LEWICKI, M. S. A review of methods for spike sorting: the detection and
classification of neural actions potentials.
Network Comput. Neural System,
v. 9, R53-R78, July, 1998.
66 LITT, B.; ECHAUZ, J. Prediction of Epileptic Seizures.
The Lancet
Neurology
, v.1, n.1, p. 21-30, 2002.
67 LITT, B.; D’ALESSANDRO, M.; et al. Translating seizure detection,
prediction and brain stimulation into implantable devices for epilepsy.
Proc. 1
st
Int. IEEE EMBS Conf. On Neural Engineering
, Capri Island, Italy, p. 485-
492, March 2003.
68 MACHADO, A. B. M.
Neuroanatomia Funcional. 2ª ed., São Paulo: Editora
Atheneu, 2003.
69 MARMARELIS, P.Z.; MARMARELIS, V.Z.
Analysis of Physiological
Systems – the white-noise approach.
New York: Plenum Press, 245p., 1978.
70 MARMARELIS, V.Z. (ed.).
Advanced Methods of Physiological Systems
Modelling.
New York: Plenum Press, v.3, 123 p., 1994.
71 MARQUES, P. A. C.
Introdução à filtragem Adaptativa. Apostila do
Professor Adjunto do DEETC do ISEL. Lisboa, Portugal. Disponível em:
<
http://www.deetc.isel.ipl.pt/Analisedesinai/pdsii/Blibliografia/Filtragem%
20Adaptativa.pdf>. Acessado em: 19 mar. 2005.
72 MELLSEN, W. J.; EPPING, W. J. M. Detection and estimation of neural
connectivity based on cross-correlation analysis,
Biological Cybernetics, v. 57,
p. 403-414, 1987.
73 MIN, L.L.; SANDER, J.W.A.S. National demonstration project on epilepsy in
Brazil.
Arch. Neuro-Psychiatr., v. 61, n. 1, p. 153-156, Mar. 2003.
169
74 MOGILNER, AY; BENABID, A-L; REZAI, AR. Brain stimulation: current
applications and future prospects.
Thalamaus Relat. Syst., v. I, p. 255-267,
2001.
75 MORMANN, F.; et al. Epileptic seizures are preceded by a decrease in
synchronization.
Epilepsy Research, v. 53, issue3, p. 173-185, 2003.
76 MORMANN, F.; KREUZ, T.; RIEKE, C.; et al. On the predictability of
epileptic seizures. Clinical Neurophysiology, p.1-19, 2004.
77 NICOLELIS, M.A.L. Controlling robots with the mind.
Scientific American,
Oct., 2002.
78 NIKIAS, C.L.; PETROPULU, A.P.
Higher-Order Spectral Analysis: A
nonlinear signal processing framework
. USA: Prentice-Hall Inc., 1993.
79 NOVELLINO, A.; CHIAPPALONE, A.; et al. Behaviors from an electrically
stimulated spinal cord neuronal network cultured on microelectrode arrays.
Neurocomputing, v. 52-54, p. 661-669, 2003.
80 NUNEZ, P.L.
Neocortical Dynamics and Human EEG Rhythms, 1
st
ed.,
New York: Oxford U. Press, 1995.
81 PAPOULIS, A.
Probability Random Variables and Stochastic Processes.
New York: McGraw-Hill, 1990.
82 PARKINSON, B. W.; SPILKER, J. J.
Global Positioning system: Theory and
Applications
, v. 163, American Institute of Aeronautics and Astronautics,
1995.
83 PEIXOTO, N.
Matrizes Neuroeletrônicas. 2001. 155f. Tese (Doutorado).
Universidade de São Paulo, São Paulo, 2001.
84 PEIXOTO, N.; MAVOORI, J.; et al. Portable Stand-alone Instrumentation for
Intracellular in Vitro Recordings.
Second International IEEE-EMBS Special
Topic Conference on Microtechnologies in Medicine & Biology
, p. 476-480,
2002.
85 PEREIRA, W.F.
Perspectivas Clínico-Terapêuticas baseadas na Engenharia
Neural
. 2004. 246f. Tese (Doutorado). Universidade Federal de Uberlândia,
Uberlândia, 2004.
86 PEYMAN, G.; et al. Subretinal semiconductor microphotodiode array.
Ophthalmic Surg. Lasers, 29, p. 234-241, 1998.
87 PFURTSCHELLER, G.; NEUPER, C. Motor imagery and direct brain-
computer interfaces.
Proc. IEEE, v. 89, p. 1123-1135, 2001.
170
88 PINE, J. Recording action potentials from cultured neurons with extracelular
microcircuit electrode.
J. Neuroscience Methods, v.2, p. 19-31, 1980.
89 Proc. IEEE.
Proceedings of IEEE, número especial sobre Engenharia Neural,
Julho, 2001.
90 RAUCH, A.; CAMERA La G.; et al. Neocortical Pyramidal Cells Respond as
Integrate-and-Fire Neurons to In Vivo-Like Input Currents.
J.
Neurophysiologic
, v.90, p. 1598-1612. Sept. 2003.
91 RENNAKER, R.L.; RUYLE, A.M.; et al. An economical multi-channel cortical
electrode array for extended periods of recording during behavior.
J. of
Neuroscience Methods
, 142, p. 97-105, 2005.
92 RIKE, F.; et al.
Spikes: exploring the neural code. New York: The MIT Press,
234 p., 1997.
93 ROUSCHE, P.; et al. “Neurabotics”: an integration of neurons, rats and robots
for advanced studies of brain-computer interfaces systems.
Proceedings of the
1
st
International IEEE EMBS, Conference on Neural Engineering, Italy,
March 2003.
94 RUTTEN, W.L.C.; SMIT, Jos P.A.; et al. Neuro-Electronic Interfacing with
Multielectrode Arrays.
IEEE Engineering in Medicine and Biology
Magazine
, v. 18, n.3, p. 47-55, 1999.
95 RUTTEN, W.L.C. Selective Electrical Interfaces with the Nervous System.
Annu. Rev. Biomed. Eng., v.4, p.407-452, 2002.
96 SAAB, M. E.; GOTMAN, J. A system to detect the onset of epileptic seizures
in scalp EEG.
Clinical Neurophysiology, v. 116, Issue 2, p. 427-442, February-
February 2005.
97 SAMAR, V.J.; et al. Wavelet analysis of neuroelectric waveforms: a conceptual
tutorial.
Brain and Language, v. 66, p. 7-10, 1999.
98 SCHLOSSHAUER, B.; et al. Towards micro electrode implants: in vitro
guidance of rat spinal cord neurites through polyimide sieves by Schwann cells.
Brain Research, 903, p. 237-241, 2001.
99 SCHUBERT, D.; HUMPHREYS, S.; BARONI, C.; COHN, M. In vitro
differentiation of a mouse neuroblastoma.
Proc. Nat. Acad. Sci., v. 64, p. 316-
323, 1969.
100 SHAHAF, G.; MAROM, S. Learning in networks of cortical neurons.
J.
Neuroscience
, v.21, p.8782-8788, 2001.
101 SHOUVAL, H. Z.; KALANTZIS, G. Stochastic Proprieties of Synaptic

171
Transmission Affect the Shape of Spike Time-Dependent Plasticity Curves.
J
Neurophysiology
, vol. 93, p. 1069-1073, Feb. 2005.
102 SILVA, G.E.G.e; COSTA NETO, T.T. da. O EEG nas epilepsias.
Jornal da
Liga Brasileira de Epilepsia
, vol.1, n.3, p. 6-9, 1998.
103 SONG, Y.; et al. Development of a Chipscale integrated
microelectrode/microelectronic device for brain implantable neuroengineering
applications.
IEEE Transactions on Neural Systems and Rehabilitation
Engineering
, v.13, n.2, p. 220-226, June 2005.
104 STAVEREN van, G.W.; BUITENWEG, J.R.; HEIDA, T.; RUTTEN, W.L.C.
Wave shape classification of spontaneous neuronal activity in cortical cultures
on micro-electrode arrays.
Proceedings of the Second Joint EMBS/BMES
Conference Houston
, TX, USA, p. 2010-2011, Out. 2002.
105 STENGER, D.A; MCKENNA, T.M.
Enabling Technologies for Cultured
Neural Networks
. New York: Academic Press, 1994.
106 STERIADE M., MCCORMICK DA, SEJNOWSKI TJ. Thalamocortical
oscillations in the sleeping and aroused brain.
Science v. 262, p. 679-685, 1993.
107 STETT, A.; et al. Biological application of microelectrode arrays in drug
discovery and basic research.
Anal Bional Chem, v.377, p. 486-495, 2003.
108 STIER, H.
Mechanismen des gerichteten Neuritenwachstums als Grundlage
Neuronaler Musterbildung.
Reutlingen, 1993. 106p. Tese (Doutorado).
Universidade de Hohenheim e Instituto de Medicina em Reutlingen
(Alemanha), 1993.
109 STRATIMIROVIC, Dj.; MILOSEVIC, S. BLESIC, S.; LJUBISAVLJEVIC, M.
Wavelet analysis of discharge dynamics of fusimotor neurons.
Physica A, v.
291, p. 13-23, 2001.
110 TEOLIS, A.
Computational Signal Processing with Wavelets: Applied and
numerical harmonic analysis.
Boston: Birkauser Ed., 1998.
111 THAKOR V.N.; TONG S. Advances in Quantitative Electroencephalogram
Analysis Methods
. Annu. Rev. Biomed. Eng., v.6, p. 453-495, 2004.
112 URL 1: Página oficial do College San Diego Mesa - USA. Imagens –
Neurônios. Disponível em:
<
www.sdmesa.sdccd.net/~bbrothers/IMAGES/neuron/neuron.jpg>. Acessado
em: 24 jan. 2006.
113 URL 2: Página oficial da revista “Ciência hoje das crianças”, n. 117, set 2001.
Disponível em:
<
http://cienciahoje.uol.com.br/materia/resources/images/chc/old/chc117c2.jpg>

172
Acessado em: 24 jan. 2006.
114 URL 3: Página oficial da Profa. Maria Bernadete Zanusso, Departamento de
Computação e Estatística/UFMS. Disponível em:
<
www.dct.ufms.br/~mzanusso/neuronio_eletronicos.htm>. Acessado em: 05
ago. 2005.
115 URL 4: Página oficial da revista “Cérebro e Mente”. Como funcionam as
Células Nervosas. Parte II. Disponível em: <http://
www.cerebromente.org.br/n10/fundamentos pot2.htm>. Acessado em 25 set.
2005.
116 URL 5: Página oficial do Programa de Pesquisa em Fronteira da Ciência.
Disponível em: <
http://www.hfsp.org>. Acessado em: 26 jan. 2006.
117 URL 6: Página oficial de Neuroanatomia da Universidade Federal da Bahia.
Disponível em:
<
http://www.neuroanatomia.ufba.br/imagens/Encefalo_superf_lat2_centro.gif>.
Acessado em: 28 jan. 2006.
118 URL 7: Página oficial do
Electroencefalograma. Disponível em:
<
www.psyvolution.pt/artigos/Electroencefalograma.pdf> . Acessado em: 28
ago. 2005.
119 URL 8: Página oficial da revista “Cérebro e Mente”. Comunicação entre as
Células Nervosas. Disponível em:
<
http://www.cerebromente.org.br/n12/fundamentos/neurotransmissores/
neurotransmitters2_p.html> . Acessado em: 25 out. 2005.
120 URL 9: Página oficial do Instituto de Ciências Biológicas da Universidade
Federal de Minas Gerais. Disponível em:
<
http://www.icb.ufmg.br/~neurofib/NeuroMed/Seminario/Epilepsia/
eletroen.htm> . Acessado em: 20 dez. 2005.
121 URL 10: ILAE – INTERNACIONAL League Against Epilepsy – Global
Campaign: What is Epilepsy? Disponível em:
<
http://www.ilae-epilepsy.org/programs/whatis.cfm>. Acessado em: 21 set.
2005.
122 URL 11: Página oficial do banco de dados de EEG, ligado ao Laboratório de
Processamento de Sinais, Universidade de Tampere - Finlândia. Disponível em:
<
ftp://sigftp.cs.tut.fi/pub/eeg-data/>. Acessado em: 10 ago. 2003.
123 VAN PELT, Jaap; WOLTERS, Pieter S.; CORNER, Michael A.; RUTTEN,
Wim L. C.; RAMAKERS, Ger J. A. Long-Term Characterization of Firing
Dynamics of Spontaneous Bursts in Cultured Neural Networks.
IEEE
Transactions on Biomedical Engineering
, v. 51, n. 11, p. 2051-2062, Nov.
2004.

173
124 VERAART, C.; RAFTOPOULOS, C.; MORTIMER, JT; et al. Visual
sensations produced by optic nerve stimulation using an implanted self-sizing
spiral cuff electrode.
Brain Res., v. 813, p. 181-186, 1998.
125 VILELA, Ana Luisa Miranda. Sistema Nervoso. In:
Anatomia & Fisiologia
Humanas
. Disponível em: <www.afh.bio.br/basicos/Nervoso1.htm>.
Acessado em: 05 maio 2005.
126 ZRENNER, E.; et al. Can subretinal microphotodiodes successfully replace
degenerated photoreceptors?
Vis. Res. 39, p. 2555-2567, 1999.
127 WEIS, R., MULLER, B., FROMHERZ, P. Neuron adhesion on a silicon chip
probed by an array of field-effect transistors.
Phys. Rev. Letters, v.76, n. 2, p.
327-330, 1996.
128 WIDROW, B.; STEARNS, S.
Adaptive Signal Processing. USA: Prentice-
Hall, 1985.
129 WOLPAW, JR.; BIRBAUMER, N.; et al. Brain-computer interfaces for
communication and control.
Clinical Neurophysiology, v.113, n.6, p. 767-791,
June 2002.
174
ANEXOS
PUBLICAÇÕES

Livros Grátis
( http://www.livrosgratis.com.br )
Milhares de Livros para Download:
Baixar livros de Administração
Baixar livros de Agronomia
Baixar livros de Arquitetura
Baixar livros de Artes
Baixar livros de Astronomia
Baixar livros de Biologia Geral
Baixar livros de Ciência da Computação
Baixar livros de Ciência da Informação
Baixar livros de Ciência Política
Baixar livros de Ciências da Saúde
Baixar livros de Comunicação
Baixar livros do Conselho Nacional de Educação - CNE
Baixar livros de Defesa civil
Baixar livros de Direito
Baixar livros de Direitos humanos
Baixar livros de Economia
Baixar livros de Economia Doméstica
Baixar livros de Educação
Baixar livros de Educação - Trânsito
Baixar livros de Educação Física
Baixar livros de Engenharia Aeroespacial
Baixar livros de Farmácia
Baixar livros de Filosofia
Baixar livros de Física
Baixar livros de Geociências
Baixar livros de Geografia
Baixar livros de História
Baixar livros de Línguas

Baixar livros de Literatura
Baixar livros de Literatura de Cordel
Baixar livros de Literatura Infantil
Baixar livros de Matemática
Baixar livros de Medicina
Baixar livros de Medicina Veterinária
Baixar livros de Meio Ambiente
Baixar livros de Meteorologia
Baixar Monografias e TCC
Baixar livros Multidisciplinar
Baixar livros de Música
Baixar livros de Psicologia
Baixar livros de Química
Baixar livros de Saúde Coletiva
Baixar livros de Serviço Social
Baixar livros de Sociologia
Baixar livros de Teologia
Baixar livros de Trabalho
Baixar livros de Turismo

{kind=link}
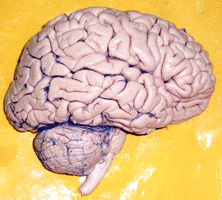{kind=link}